
基本信息
編號:S00013;
學名: Heterodera avenae Wollenweber,1924;
 燕麥胞囊線蟲病
燕麥胞囊線蟲病異名:Heterodera schachtii var. avenae Wollenweber,1924;H. schachtii subsp maior O.schmidt,1930;
H. maior(O.schmidt,1930)Franklin,1940 H.ustinovi Kirjanova,1969;
Bidera ustinovi Kirjanova,1969;
Krall & krall,1978 H.B. avenae(Wollenweber,1924) Krall & krall,1978;
寄主植物
寄主範圍比較窄、燕麥胞囊線蟲的模式寄主是燕麥(Avena sativa)、最主要的寄主是早熟禾(Poaceae)和禾本科(Gramineae)、包括經濟上最重要的禾穀類:燕麥(Avena)、小麥(Triticum)、大麥(Hordeum)、以及黑麥(Secale)、冰草屬(Agropyrum)、剪谷穎(Agrostis)、看麥娘(Alopecurus)、短柄草屬(Brachypodum)、雀麥屬(Bromus)、洋狗尾草(Cynosurus)、稗草屬(Echimochloa)、羊茅(牛尾)草屬(Festuca)、洽草屬(Koeleria)、毒麥屬(Lolium)、草屬(Phalaris)、梯牧草屬(Phleum)、早熟禾屬(Poa)、棒頭草屬(Polypogum)、狗尾草屬(Setaria)、高粱屬(Sorghum)、和玉蜀黍(Zea)(Bovien、1953;Winslow、1954;Duggan、1958;Hesling、1958;stone、1960;Gill及Swarup、1971)、也有一些寄主不是禾穀類植物、包括許多雜草、儘管不是好的寄主、但這些雜草仍可維持穀類作物間的線蟲種群。
分布情況
燕麥胞囊線蟲是世界性分布的重要病原線蟲,中國境外:英國、前蘇聯、義大利、澳洲、美國、加拿大、日本和印度,已在五大洲的32個產麥國家發生為害。
中國境內:湖北天門縣、潛江、仙桃、新洲、河北、河南、山西的太谷、安徽的固鎮、北京市近郊。
為害症狀
線蟲危害寄主的地上部分症狀並不專化,在田間,麥類受燕麥胞囊線蟲為害後早期即有表現,植物出現小塊面積黃化,生長衰弱矮化。受到嚴重侵染的燕麥幼苗的葉片可伴有少量蟲癭。
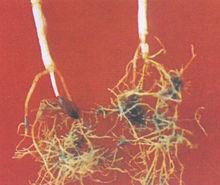
寄主的根系受害症狀,小麥播種後待幼苗生長3~6周時,小心挖出小麥根系,可看到受線蟲嚴重侵染的植株形成多結的根系。根係數量減少,高度分枝並增生出短粗的側根。根系分枝多,細而亂,甚至糾結成團。開花前後,根側露出微小如粉末的發亮顆粒狀物(即膨大的雌成蟲或白色胞囊)。胞囊直徑小於1mm,易與化肥或石灰石顆粒及沙粒相混淆。與絲核菌(Rhizoctona solani)混發,可加重一些症狀的表現。
測量數據
雌蟲(♀♀):L=0.55~0.77mm,W=0.36~0.50mm。
雄蟲(♂♂):L=1.07~1.59(1.38)mm,a=32~55(45),b=7~12(9),bˊ=5.5~6.8(5.8),T=40~50(45),口針長=27~31(28.5μm),引帶長=33~38(36)μm。
2齡幼蟲:L=0.52~0.61mm(大多數在0.54~0.58mm範圍內,平均0.575,據Franklin,1951),寬=20~24μm,尾長=45~70μm(通常54~58μm),口針長=24~28μm(平均27μm,據Triffitt,1929)。
形態描述
雌蟲:成蟲檸檬形,頸部和陰門錐明顯,唇區有環紋。6個唇瓣會合,有一個唇盤;口針直或略呈弓形,26~36μm長,口針基球圓。中食道球圓形且有明顯的瓣門。陰門裂12~13μm長,陰門處有膠質狀的擠壓物,但擠壓時很少有卵。角質表面有鋸齒形的皺縮的紋飾。亞晶層明顯,在胞囊呈暗綠色時脫去。胞囊的陰門錐大部分被無色透明的陰道結構占滿;泡狀突(bullae)明顯且聚集在陰門錐下,雙半膜孔。
胞囊:平均大小為0.71mm×0.50mm(Triffitt,1929)。成熟胞囊由白色變為褐色時不經歷黃色階段(Franklin,1951)。
卵:從新模式標本採集地(Aschersleben,德國)采來的20個卵的平均值:長12μm,寬56μm。
雄蟲:蠕蟲形,數量多。體環紋明顯,大約2μm寬。側區有4條側線,外部帶狀區通常呈網紋狀。唇區圓,縊縮,有4~6條環紋(通常5條)。頭架深度骨化,有明顯的外緣。頭小體不明顯,位於唇區後前部1~2個體環及後部6~7個體環處。口針前部的錐體部分迅速變尖,通常短於口針長度的一半。口針基球圓形,通常有一平的向後傾斜的前表面;背食道腺開口在口針基球後3~6μm處。中食道球橢圓形,具明顯瓣門。食道覆蓋腸的腹側和側腹;排泄孔在食道與腸連線處附近。半月體明顯,2~3個體環長。通常位於排泄孔前5~6個體環處,但有時僅在其前1~2個體環處。半月小體不明顯,在排泄孔後6~7個體環處。交合刺弓形,腹面有一中等大小的凸緣和一具缺刻的末端,兩者形成一長的窄管。引帶簡單,稍彎曲。尾末端通常捲曲。
2齡幼蟲:蠕蟲形,尾端聚尖。唇區圓,縊縮,有2~4條環紋。體環明顯,近中部平均1.5μm寬。側區寬約為體寬的1/4,有4條側線並形成3條帶,外側帶網紋形。口針發達,口針基球大,其前部扁平或凹陷;前部錐體不到口針長度的一半。中食道球圓,肌肉質發達,具有一大的瓣門。尾長為肛門處體寬的3~4.5倍;無色透明區部分長35~45μm或約為1.5個口針長。側尾腺孔形,明顯,恰好位於肛門後。
生活史介紹
線蟲從卵孵化出2齡幼蟲,侵入寄主根系,繼續發育,蛻皮3次,變為成蟲,雌雄交配,雌蟲產卵,完成發育循環,這就是線蟲的生活史。
生物學特性
1、線蟲的生活史
線蟲從卵孵化出2齡幼蟲,侵入寄主根系,繼續發育,蛻皮3次,變為成蟲,雌雄交配,雌蟲產卵,完成發育循環,這就是線蟲的生活史。
H.avenae可在世界範圍內很廣的生態環境中發生,在一年內僅發生1代或2代。
2、線蟲的寄生性
燕麥胞囊線蟲種記憶體在生理分化現象,國際已正式命名13個致病類型,歐洲的致病型與澳大利亞、印度的致病型不同。鄭經武等(1997)套用國際鑑別寄主A組11個品種,鑑定中國山西太谷和安徽固鎮的兩個燕麥胞囊線蟲群體的致病型,結果指出,我國這兩個地區的基本屬於一個致病型,同時與國外的13個致病型不同,治病譜不寬,而與瑞典的致病型相似。
3、生活習性
卵可在胞囊記憶體活數年,尤其是在涼爽、乾燥的條件下存活時間更長。在田間條件下,經歷一段潮濕、涼爽的階段後,卵開始孵化,卵孵化要求的溫度條件比較複雜,但涼爽的氣候條件(2~10℃)是卵孵化的最適溫度。白色雌蟲的陰門內可分泌出膠狀的物質,但卵並不從陰門裂產出。胞囊內的卵孵化出2齡幼蟲。侵染性幼蟲(J2)進入根冠後部並在細胞內移動,並在內皮層內逐漸使頭部與主軸保持平行。可在內皮層和中柱鞘附近建立取食位點。侵入後6~9周內雌蟲成熟,而雄蟲在3周內離開根系。在秋季和初冬播種穀類作物,麥類萌芽生根時則正好與本年的侵染時期相遇。2齡幼蟲可侵入根系,越冬,但不繼續發育,次年春季繼續發育。春播區,在歐洲大約在3月中旬至6月中旬2齡幼蟲陸續從胞囊孵化出來並侵入根系。在澳大利亞大量胞囊出現在8月末和9月,遲的可在10月份出現(Meahger,1968);而在印度的拉賈斯坦在2月份出現胞囊(Singh & Swarup,1964)。不管在何處發生的H.avenae,每年是否能完成一代以上仍不清楚。除去寄生作物之外,線蟲的分布還與土質有關。在英國,燕麥胞囊線蟲在輕砂性土、貧瘠土壤和缺少有機質的白堊土中分布最多,為害也最重。
王明祖(1993)取湖北天門的病土,在武漢做孵化試驗,以旬平均溫度9~12℃孵化出的幼蟲最多。在15℃(±)1℃恆溫下,孵化數量高於在20℃,10℃,5℃下的孵化數。在自然情況下,6~10月無線蟲孵化。晾乾的胞囊在冰櫃冷凍室記憶體放12個月不影響孵化。劉景瑞等報告(1995)在河南省安陽地區一年發生一代,侵染期再3月中旬以前,5月中旬以後白色的胞囊逐漸增多,在6月上旬達到高峰。盆栽試驗證明在秋季不能侵染小麥。
張東升等(1994)室外盆栽試驗證明,在北京地區在冬小麥上有很高的繁殖率。王振躍報告,1991~1992年調查,燕麥胞囊線蟲在鄭州郊區發生,人工接種小麥、大麥、燕麥和黑麥是很好的寄主,而穀子、玉米、水稻和高粱接種後不顯症狀,在根內未查到線蟲。
傳播途徑
胞囊失去生命後脫落入土中越冬,可借水流、風,農機具等傳播。
檢疫方法
1、受檢材料 這種線蟲休眠和被攜帶的形態是胞囊,而胞囊主要存在於土壤中或混在附有土壤的禾穀類植物的種子、根系等材料上,上述進境材料均屬受檢範圍。
2、採樣 上述進境材料,隨機取樣待檢;對於已經播種的地塊,在播種後幼苗長到3~6周時,小心挖出根系,檢查根系是否被侵染。在作物開花期可檢查根上是否有白色雌蟲,採集根際土,分離胞囊。
3、分離方法 見其他胞囊線蟲的分離方法。
防治方法
1、植物檢疫
我國過去一直未見發生燕麥胞囊線蟲的報告,目前僅是少數地區小面積發生,劃為疫區,全力撲滅。實行嚴格的檢疫制度,限制H.avenae向無病區擴散與傳播,這是全國麥區的迫切任務。
2、農業栽培技術措施
(1)輪作 通過與非寄主植物(如豆科植物大豆、豌豆、三葉草和苜蓿)和不適合的寄主作物輪作,可降低H.avenae的田間群體密度。而與非寄主作物—如豆科植物進行3年的輪作可使線蟲數量大為降低,而且這樣的輪作是經濟有效且可行的。在澳大利亞,與豆科植物進行輪作的另一個好處是能改善土壤氮素缺乏的狀況。
在我國病區實行與水稻、油菜、棉花等輪作是切實可行的。
(2)調節播期 土壤溫度對H.avenae的生活史及其對寄主的為害有著極大的影響,低溫可以刺激卵內幼蟲的孵化和抑制寄主根系的生長。在維多利亞和澳大利亞,提倡適時播種,一般在5月下旬土壤溫度降至8~12℃時就播種,隨著溫度再降低幼蟲大量孵化侵染時,小麥植株的根系已發育良好抗侵染能力增強,降低了線蟲的為害程度。
3、抗病品種
我國發生地區沒有對燕麥胞囊線蟲的小麥抗病品種,據相關數據顯示,是加強種質資源的鑑定,尋找抗源,選育抗病品種。
根據國外的經驗,套用抗病品種,對大麥和燕麥來講,這項措施對防治H.avenae很適用,但因為線蟲存在一些專化性的致病變異類型,所以僅在一定的環境條件下這一措施才有效。與大麥和燕麥相比,小麥對H.avenae具有抗性的抗源相對較少。在大麥和燕麥中,抗性基因通常不與不良性狀相連鎖,但小麥的抗性基因是有限的,儘管在硬粒小麥中一些品系是有效的。O’Brien(1972)證明小麥品種(Aus10894)和(Loros)中有一個相同的顯性基因位於小麥B染色體組的第2條染色體上,對澳大利亞的燕麥胞囊線蟲的致病型Ha13表現抗病。Meleod(1973)套用Aus10894和Festiguay已選育出了Katyil、Molineux和Kellalac等對致病型Ha13表現抗病的品種。East Wood(1991)在小麥的近緣種中尋找抗病基因,發現節節麥(Triticum tauschii)是燕麥胞囊線蟲的新抗源。利用此抗源已選育出了一系列的抗病品種和品系—Aus 18912、Aus 19813,CPI 110809,CPI 110810和Aus 18914等。
4、化學防治
過去,在燕麥胞囊線蟲嚴重發生的地塊,可用D-D混劑、氯化苦及棉隆(dazomet或Basamid、Mylone)、涕滅威*(Temik或aldicarb)有效地進行防治。但在禾穀類作物田間進行化學防治是不經濟的,也不實用,殺線劑嚴重污染環境。
5、生物防治
生物防治在防治燕麥胞囊線蟲上很有發展前景,一些生物因子,尤其一些真菌對抑制燕麥胞囊線蟲的種群很有效。在北歐曾發現連續多年種植禾穀類作物後,田間燕麥胞囊線蟲的群體反而下降,即使播種感病的小麥品種受害的程度也減輕,後來證明主要是嗜線疫霉(Nematophthora gyhophila)和(Pochonia chlamydosporia)(Verticillium chlamydosporium)的作用,兩種真菌寄生雌蟲和卵。在澳大利亞和維多利亞西部的麥田中,V. chlamydosporium的分布非常普遍,此外還發現了Caenaria auxiliaris、Hirsutella rhossiliensis和Nematoctonus haptocladus等多種寄生菌。這些寄生菌的自然寄生,一定程度上能降低田間有生活力卵的數量。和許多其他的植物寄生線蟲的生物防治一樣,燕麥胞囊線蟲的生物防治也還處在試驗研究階段。
