
釋義
棲地碎裂化(habitat fragmentation)是指在自然干擾或人為活動的影響下,大面積連續分布的棲息地被分隔成小面積不連續的棲息地斑塊的過程。這種干擾可以形成多種空間模式,從棲息地被小範圍的中斷到殘餘斑塊在已經轉變了的基質中的零星散布,都屬於棲地碎裂化。因此,棲地碎裂化既可以理解為棲息地斑塊的空間模式,也可以理解為產生這種模式的過程。最初的棲地碎裂化概念既包括原棲息地面積的喪失也包括棲息地空間格局的動態變化,近年來,多數學者主張將棲息地喪失與空間格局的變化這兩個概念分開,主要原因是棲息地喪失與空間格局變化的物理結果不一樣,並且二者對野生動物的生態學效應存在著一定的差異,棲地碎裂化概念用於特指棲息地空間格局的改變。
後果及影響
保護生物學家和生態學家普遍認為棲地碎裂化有各種各樣的消極後果。棲地碎裂化除了縮小原有棲息地的總面積外,棲息地斑塊的面積也會逐漸減少,致使棲息地斑塊廣泛的分離,臨近邊緣的棲息地比例增加,邊緣也變得越來越分明。由於面積效應的作用,致使野生動物的種群數量減少,最終導致某些種類在小面積的斑塊中消失,同時還可增加棲息地斑塊中種群對干擾的敏感性。由於棲息地斑塊的孤立和隔離,致使局部滅絕後的重新建群變得緩慢。有些物種如大型捕食者和留鳥對這些效應的高度敏感性會導致物種多樣性的減少和群落結構的變化。由於邊緣效應的作用,殘餘森林斑塊內的種群和群落動態受捕食、寄生和物理干擾等因素的控制。另外,這些變化通常伴隨著潛在的間接效應。例如,棲地碎裂化如果影響了昆蟲的分布和豐富度,食蟲鳥的資源基礎就會發生改變。
棲地碎裂化過程
棲地碎裂化是一動態過程
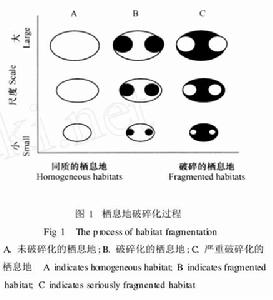 棲地碎裂化
棲地碎裂化棲息地的破碎化可定義為對連續性棲息地的擾動過程,而這種擾動又可產生一系列的空間格局,無論是過程還是格局都處於動態之中。在理解棲地碎裂化時首先要界定什麼樣的棲息地是破碎的和什麼樣的棲息地不是破碎的,右圖描述了棲地碎裂化過程及其不同階段的空間格局,可以從圖中看出哪些棲息地是未破碎的、哪些棲息地是破碎的以及哪些棲息地是嚴重破碎的,即棲地碎裂化程度的梯度變異。以往的研究經驗告訴人們破碎化程度可以通過測量斑塊面積和隔離度而獲得,這種研究方式隨著景觀生態學的滲透理論的套用而得到補充和加強。在具體問題研究過程中,往往將未破碎的棲息地稱為同質的棲息地,並於尺度關係密切,棲地碎裂化格局隨著研究尺度的改變而變化。圖中所描述的梯度是連續的,而不是單獨的以隔離度為基礎任意地分為“已破碎的”和“未破碎的”,因為棲地碎裂化是一連續的動態過程。
動物種群對棲地碎裂化過程的不同階段有著不同的反應,即動物種群對棲地碎裂化的反應受時間動態的影響。在棲地碎裂化初期,由於對某一地段的習慣,有些個體對棲息地的變化不會立即做出反應,這一現象容易隱藏和掩蓋破碎化效應,從而對某些動物種群形成“生態陷阱”。另外,在棲地碎裂化過程中,個體間的替代作用和種群中的漂泊者會產生類似的效果。有些物種對棲地碎裂化的反應是快速地進入殘餘的棲息地斑塊中,所以殘餘的棲息地斑塊中暫時增加了物種豐富度和個體密度。在破碎化棲息地中,群體的社會吸引力也能改變個體的分布模式從而在對棲息地變化的反應上產生時滯。然而,隨著棲地碎裂化過程的持續和時間序列的變化,破碎化效應對動物種群的各種不良後果會相繼出現。
尺度的重要性
空間和時間尺度包含於任何生態系統的過程中,對於自然現象的描述與選擇的尺度有很大關係,在不同尺度上,生態系統模式的表現過程往往不一致。例如,在地中海區域,有些鳥類的空間分布格局在細小尺度上可以表現為均勻型而在較大尺度上又表現為聚集型。尺度越大,生態系統的成分就越複雜,但其變異性往往隨著尺度的增大而縮小,可預測性增強。相反,小尺度下的生態系統儘管成分相對簡單,但由於隨機現象的增加,使生態系統變得不可預測。VanTurnhout等在不同空間尺度上對荷蘭的繁殖鳥類多樣性進行了25a的監測,發現在不同尺度下,鳥類物種多樣性的表現模式並不一樣,他們的實驗結果證實了在較大尺度上鳥類多樣性的分布模式具有更強的可預測性。
尺度的變化可能會影響人們對棲地碎裂化的理解,不應該試圖決定哪一種尺度是最合理的,而應該把工作重點放在理解和認識研究對象在不同尺度上的發生規律。最佳生態尺度的選擇隨著研究對象和研究問題的不同而發生變化。例如,一片面積為100hm森林破碎成10個森林斑塊,對領域面積相對較小的白喉林鶯來說,必須面臨著一種選擇,即正確的判斷哪些斑塊是適合生存的,哪些不是,如果判斷有誤,可能帶來滅頂之災。而這種破碎化對領域面積較大蒼鷹似乎不產生影響,這10個斑塊只是蒼鷹棲息地中細密的紋理。生態尺度的選擇還與研究的焦點是動物個體還是動物種群密切相關。對於同一種鳥類而言,研究種群的尺度往往大於研究個體的尺度。例如,一隻雄性松雞在繁殖季節需要20~50hm的森林作為領域,一個松雞繁殖種群往往需要10000hm的林地作為繁殖場所。另外,同一物種在不同的區域領域面積差異較大,在選擇研究尺度時應該加以區別,例如,斑點林鴞個體的年領域範圍在不同的區域從500hm到5000hm不等。
棲地碎裂化研究的理論基礎
到目前為止,有關棲地碎裂化研究的理論基礎主要有島嶼生物地理學理論、景觀生態學中的結構與功能原理和集合種群理論。在利用島嶼生物地理學理論對生境破碎化進行研究時,有兩點假設:一是將斑塊比喻成海洋中的島嶼;二是假設斑塊周圍的異質環境對斑塊中的有機體來說是難易逾越的“海洋”。然而,事實並非如此。棲息地斑塊之間的通透性遠遠大於真正的島嶼。斑塊動物種群受周圍環境的影響較大,這些影響有時甚至大於斑塊內部的結構效應。所以,島嶼生物地里理論在研究棲地碎裂化時受到了一定的局限。景觀生態學的結構與功能原理從複雜的空間格局出發,將空間格局定義為基質、斑塊和廊道等元素來研究破碎生境中的種群動態,充分考慮了生境的空間異質性。結構與功能原理強調環境的複雜物理結構,探求不同空間尺度上異質物理環境所產生的種群、群落與生態系統水平上的生態後果,由於這種複雜性,所以數學建模和量化分析困難重重。集合種群理論充分考慮了斑塊空間格局對種群的影響,對空間環境的簡化使其理論內涵與實際套用能有效地結合在一起。經典的集合種群理論由Levins於1969年提出,在隨後的幾十年里McCullough、Schwartz、Hanski等學者對該理論進行了完善和補充,提出了狀態轉換模型、多種群模擬模型和關聯函式模型等。Pulliam及Pulliam和Danielson提出了源種群、匯種群以及相應的源生境、匯生境的概念,為集合種群理論增添了新的活力,並使集合種群理論得以廣泛地套用到棲地碎裂化對動物種群影響的研究中。
