
基本介紹
為保持生態平衡,保護生物資源,發展畜牧業、漁業,防治病蟲害,都需要研究生物種群的變化和發展,特別是種群的規模。所謂種群是指在特定的時間裡占據一定空間的同一物種的有機體的集合,影響種群動態變化的原因是很多的,至今還往往不能說清究竟哪些因子影響著所觀測種群的變化,因此我們所得到的模型經常是比較粗糙的,但這些模型的討論能很好地展示數學建模的過程,對了解數學建模方法的實質是有幫助的。
運用數學方法研究種群的變化規律,通常是根據大量的實驗或統計資料,作出某些假設,選取有關變數,確定有關參數,建立數學方程,然後利用數學的理論與方法進行分析,去揭示種群的變化規律,預測它的未來,再通過實踐去檢驗和不斷修正、深化原有模型。
生物種群模型可分為 單種群模型和多種群模型,利用生物種群模型主要是研究當前時刻某種群的數量、預測未來某一時刻種群的數量和預測生態平衡時種群的數量 。
種群的幾何級數增長
種群的幾何級數增長(geometric growth)是指種群在無限的環境中生長,不受食物、空間等條件的限制,種群的壽命只有1年,且一年只有一個繁殖季節,同時種群無年齡結構,彼此隔離的一種增長方式。假設有一理想種群,開始時有10個個體,且每個個體一年繁殖1次,每次產生2個後代,則到第2代時,種群個體將上升為20個,以後每代增加1倍,依次為40,80,160…一這種現象可用如下的簡單公式描述。即:
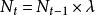 單種群模型
單種群模型或
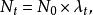 單種群模型
單種群模型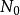 單種群模型
單種群模型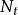 單種群模型
單種群模型式中: 為初始種群大小, 為時間t時的種群大小,λ是種群的周期增長率。
根據以上模型可以計算世代不相重疊種群的增長情況。即當λ>1時,表示種群增長;λ=1時,種群穩定:λ<1時,種群下降,當λ=0時,種群無繁殖現象,且在下一代滅亡 。
(指數型增長)Malthus模型
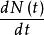 單種群模型
單種群模型在無限環境條件下,除了種群的離散增長外,有些生物可以連續進行繁殖,沒有特定的繁殖期,在這種情況下,種群的增長表現為指數形式。種群的增長被認為是一個連續過程,並且環境容納無限大,設N(t)為時刻t時種群的數量,則種群的瞬時增長率 可以下式表達
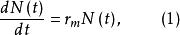 單種群模型 單種群模型
單種群模型 單種群模型這裡,r為種群的內稟增長率,其值等於出生率減去死亡率,如果在時間t=0時種群的大小為 ,那么式(1)的解為
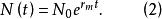 單種群模型
單種群模型內稟增長率是指在環境條件無限制作用時,由種群內在因素決定的最大相對增殖速度,其單位為時間的倒數。對某一種群來說,內稟增長率是當種群建立了穩定的年齡分布時,某穩定的相對增長率,它反映的是一種理想狀態,可用來與實際條件下的增長率進行比較,其差值可視為環境對生物增殖阻力的量度,稱為環境阻力。
以觀測的種群數量與時間t做圖,種群增長曲線呈“J”字型,故指數增長又稱J增長。
具有指數增長特點的種群,其數量變化與r值關係密切。當r>0時,種群數量呈指數上升;r=0時,種群數量不變;r<0時,種群數量呈指數下降。圖1給出了4個不同r值的種群增長曲線,其中有兩個r值大於零,一個小於零 。
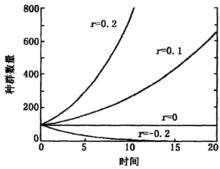 圖1 4個不同r值的種群增長曲線(種群原始數量為100)
圖1 4個不同r值的種群增長曲線(種群原始數量為100)Logistic模型
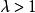 單種群模型
單種群模型從種群的指數增長模型可知,只要 或r值大於零,種群就會持續增長,即形成無限增長,但在實際環境下,由於種群數量總會受到食物、空間和其他資源的限制,因此,增長是有限的。由於環境對種群增長的限制作用是逐漸增加的,故增長曲線呈現“S”型,也稱S型增長,其數學模型可用logistic方程描述 :
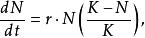 單種群模型
單種群模型式中:N為種群數量,K為環境容量(earring capaity)。
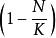 單種群模型
單種群模型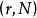 單種群模型
單種群模型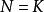 單種群模型
單種群模型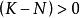 單種群模型
單種群模型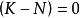 單種群模型
單種群模型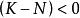 單種群模型
單種群模型即某一環境所能維持的種群數量,在曲線中表示為漸近線。當N由0增加到K時, 則由1變化到0,即隨種群數量N的增長,種群的指數增長 實現的程度逐漸變小,直到 時,增長為0。故當 時,種群增長; 時,種群停止增長; 時,種群數量下降。
真實的種群不可能有無限大的環境容量,種群的增長總存在一個上限K,當種群的數量N(t)逐漸向其上限K值增加時,實際增長率就要逐漸減少,我們有:
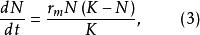 單種群模型
單種群模型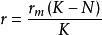 單種群模型
單種群模型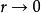 單種群模型 單種群模型
單種群模型 單種群模型式(3)中,實際增長率為 ,當種群大小到達K值時, ,表明增長率r與種群密度問成反比的關係,稱為對增長率的密度制約效應。如果在t=0時種群大小為 ,則式(3)的解為:
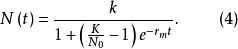 單種群模型
單種群模型邏輯斯諦曲線常劃分為5個時期:①開始期,也可稱潛伏期,由於種群個體數量少,密度增長緩慢;②加速期,隨個體數的增加,密度增長逐漸加快;③轉折期,當個體數達到飽和密度一半(即K/2)時,密度增長最快;④減速期,即個體數超過K/2以後,密度增長逐漸變慢;⑤飽和期,種群個體數達到K值而飽和。
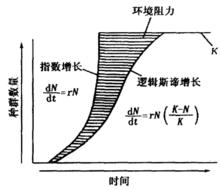 圖2 種群增長模型
圖2 種群增長模型邏輯斯諦方程的兩個參數r和K均具有重要的生物學意義。如前所述,r表示物種的潛在增殖能力,而K則表示環境容納量,即物種在特定環境中的平衡密度。雖然模型中的K值是一最大值,但從生物學含義講,它應該並可以隨環境(資源量)的改變而改變。
邏輯斯諦方程的重要意義是:①它是許多相互作用種群增長模型的基礎;②它也是漁業、林業、農業等實踐領域中確定最大持續產量(maximum sustained yield)的主要模型;③模型中兩個參數r、K,已成為生物進化對策理論中的重要概念。
在農業生態系統中,不同的生物種群所遵循的增長方式是不同的。如某些細菌、昆蟲、鼠類的繁殖,基本上屬於“J”增長型,一年生植物的乾物質在某段時間內的增長也屬於“J”增長型。多數生物的增殖,包括植物分櫱的增加,株高的增長基本上屬於“S”型。有些昆蟲在環境有利的年份數量迅速增加,呈現“J”型增長;而在環境條件不利的年份則呈“S”型增長。
Logistic方程比Malthus方程更能符合實際情況。但Logistic方程仍有缺點,即沒有考慮到種群的年齡分布,對於壽命長的、世代重疊多的種群,此方程仍會產生很大的偏差。只有低級的生物,如細菌、酵母或浮遊動植物才與之比較吻合 。
有收穫的單種群模型
Logistic方程描述種群在自然環境下的增長規律。這些種群如果是某種具有經濟價值的動植物的種群,人類不時對其進行收穫,種群模型是怎樣的昵?比如在漁業中,式(3)只描述魚群在自然環境下生長的情況,沒有考慮有捕撈的情況。如果考慮到捕撈因素,則模型(3)要作相應的修改。
(1)具常收穫率的單種群模型
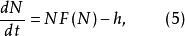 單種群模型
單種群模型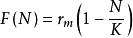 單種群模型
單種群模型其中,F(N)表示r與種群密度的關係,對於LogistiC方程而言,,h為常收穫率。
(2)具時變收穫率的單種群模型
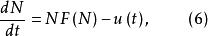 單種群模型
單種群模型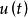 單種群模型
單種群模型為與時間有關的收穫率 。
具時滯的單種群模型
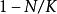 單種群模型
單種群模型Logistic模型考慮的關於r的調節因子是,它是與瞬時密度有關的調節機理。但在實際情況中,這種調節效應大多數都有某種時滯效應,即因子與效果之間差一個時間因子T,於是便得到時滯模型 :
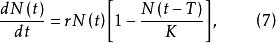 單種群模型
單種群模型式(7)表明,時刻t種群的增長不僅與時刻t的種群大小有關,而且與以此前時間T的種群密度有關。如果種群增長與過去所有時間的種群密度都有關係,這種情況下,具有時滯的種群增長模型為:
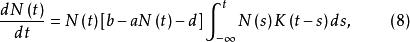 單種群模型
單種群模型這裡a、b、d均為常數。
關於一般的密度制約的情況下,有
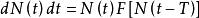 單種群模型
單種群模型或
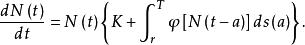 單種群模型
單種群模型其他情況
另外還有離散時間的單種群模型,如果物種世代之間沒有重疊,所以種群增長分步進行,描述它們的生長過程是一個不連續的模型,一般是一個差分方程。還有具時變環境的單種群模型等 。
