
定義
 古核生物的結構
古核生物的結構古細菌(archaeobacteria)(又可叫做古生菌、古菌、古核細胞或原細菌)是一類很特殊的細菌,多生活在極端的生態環境中。具有原核生物的某些特徵,如無核膜及內膜系統;也有真核生物的特徵,如以甲硫氨酸起始蛋白質的合成、核糖體對氯黴素不敏感、RNA聚合酶和真核細胞的相似、DNA具有內含子並結合組蛋白;此外還具有既不同於原核細胞也不同於真核細胞的特徵,如:細胞膜中的脂類是不可皂化的;細胞壁不含肽聚糖,有的以蛋白質為主,有的含雜多糖,有的類似於肽聚糖,但都不含胞壁酸、D型胺基酸和二氨基庚二酸。
生存環境及形態
生存環境
很多古菌是生存在極端環境中的。一些生存在極高的溫度(經常100℃以上)下,比如間歇泉或者海底黑煙囪中。還有的生存在很冷的環境或者高鹽、強酸或強鹼性的水中。然而也有些古菌是嗜中性的,能夠在沼澤、廢水和土壤中被發現。很多產甲烷的古菌生存在動物的消化道中,如反芻動物、白蟻或者人類。古菌通常對其它生物無害,且未知有致病古菌。
形態
單個古菌細胞直徑在0.1到15微米之間,有一些種類形成細胞團簇或者纖維,長度可達200微米。它們可有各種形狀,如球形、桿形、螺旋形、葉狀或方形。它們具有多種代謝類型。值得注意的是,鹽桿菌可以利用光能製造ATP,儘管古菌不能像其他利用光能的生物一樣利用電子鏈傳導實現光合作用。
進化和分類
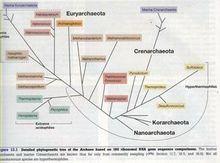
從rRNA進化樹上,古菌分為兩類,泉古菌(Crenar
 古核生物的結構
古核生物的結構chaeota)和廣古菌(Euryarchaeota)。另外未確定的兩類分別由某些環境樣品和2002年由Karl Stetter發現的奇特的物種納古菌(Nanoarchaeum equitans)構成。 Woese認為細菌、古菌和真核生物各代表了一支具有簡單遺傳機制的遠祖生物的後代。這個假說反映在了“古菌”的名稱中(希臘語archae為“古代的”)。隨後他正式稱這三支為三個域,各由幾個界組成。這種分類後來非常流行,但遠組生物這種思想本身並未被普遍接受。一些生物學家認為古菌和真核生物產生於特化的細菌。
古菌和真核生物的關係仍然是個重要問題。除掉上面所提到的相似性,很多其他遺傳樹也將二者並在一起。在一些樹中真核生物離廣古菌比離泉古菌更近,但生物膜化學的結論相反。然而,在一些細菌,(如棲熱袍菌)中發現了和古菌類似的基因,使這些關係變得複雜起來。一些人認為真核生物起源於一個古菌和細菌的融合,二者分別成為細胞核和細胞質。這解釋了很多基因上的相似性,但在解釋細胞結構上存在困難。
目前有22個古菌基因組已經完全結束了測序,另外15個的測序工作正在進行中。
代表性古細菌
極端嗜熱菌(themophiles):能生長在90℃以上的高溫環境。如史丹福大學科學家發現的古細菌,最適生長溫度為100℃,80℃以下即失活,德國的斯梯特(K. Stetter)研究組在義大利海底發現的一族古細菌,能生活在110℃以上高溫中,最適生長溫度為98℃,降至84℃即停止生長;美國的J. A. Baross發現一些從火山口中分離出的細菌可以生活在250℃的環境中。嗜熱菌的營養範圍很廣,多為異養菌,其中許多能將硫氧化以取得能量。
極端嗜鹽菌(extremehalophiles):生活在高鹽度環境中,鹽度可達25%,如死海和鹽湖中。
極端嗜酸菌(acidophiles):能生活在pH值1以下的環境中,往往也是嗜高溫菌,生活在火山地區的酸性熱水中,能氧化硫,硫酸作為代謝產物排出體外。
極端嗜鹼菌(alkaliphiles):多數生活在鹽鹼湖或鹼湖、鹼池中,生活環境pH值可達11.5以上,最適pH值8~10。
產甲烷菌(metnanogens):是嚴格厭氧的生物,能利用CO2使H2氧化,生成甲烷,同時釋放能量。 CO2+4H2→CH4+2H2O+能量
由於古細菌所棲息的環境和地球發生的早期有相似之處,如:高溫、缺氧,而且由於古細菌在結構和代謝上的特殊性,它們可能代表最古老的細菌。它們保持了古老的形態,很早就和其它細菌分手了。所以人們提出將古細菌從原核生物中分出,成為與原核生物(即真細菌eubacteria)、真核生物並列的一類。
嗜熱細菌
嗜熱細菌只有在高溫下才能良好地生長。迄今為止已分離出50多種嗜熱細菌。在這些細菌中有一種最抗熱的菌株(Phyolobous fumarii),在105℃繁殖率最高,甚至在高達113℃也能增殖。深海極端嗜熱和產甲烷細菌,備受人們關注,因為它位於生命進化系統樹的根部附近,對它進行深入研究,可能有助於我們弄清世界上最早的細胞是如何生存的問題。有人認為嗜熱細菌生存的極限溫度可能是150℃,若超過這一溫度,無論哪種生命形式都不可避免地使維持DNA和其他重要的生命大分子完整性的化學鍵遭到破壞。PCR(多聚酶鏈反應)中所使用的Taq酶就是從T.aquaticus嗜熱細菌中分離到。現又從Pyrococcus furiosus分離一種Pfu聚合酶取代了Taq酶,Pfu酶在100℃時能最好地發揮作用。
嗜鹽細菌
它能在極端地鹽環境下生長和繁殖,特別是在天然地鹽湖和太陽蒸發鹽池中生存。由滲透勢原理可知,高鹽溶液中的細胞將失去更多的水分,成為脫水細胞。而嗜鹽細菌可產生大量的內溶質或保留從外部取得溶質的方式來維持自身的生存,如嗜鹽桿菌(Halobacterium salinarum)在其細胞質內濃縮了高濃度氯化鉀,其中有一種酶只有在高濃度的氯化鉀中,才有活性,才能發揮其功能。而與環境中鹽類接觸的鹽桿菌,其細胞質中的蛋白質需要有高濃度的氯化鈉才能發揮作用。
歷史
古細菌這個概念是1977年由Carl Woese和George Fox提出的,原因是它們在16SrRNA的系統發生樹上和其它原核生物的區別。這兩組原核生物起初被定為古細菌(Archaebacteria)和真細菌(Eubacteria)兩個界或亞界。Woese認為它們是兩支根本不同的生物,於是重新命名其為古菌(Archaea)和細菌(Bacteria),這兩支和真核生物(Eukarya)一起構成了生物的三域系統。
70年代末,沃斯等人用他們獨創的技術分析了200多種細菌和真核生物(包括其中的某些細胞器)的16S(或18S)核糖體核糖核酸(rRNA)的寡核苷酸譜,結果將生物分為3大類群:真核生物、真細菌和古細菌。
古細菌包括3類不同的細菌:產甲烷細菌、極端嗜鹽細菌和嗜酸嗜熱細菌。它們生存在極端特殊的生態環境中,具有獨特的16S核糖體RNA寡核苷酸譜。而且,它們在分子水平上與真核生物和真細菌都有不同之處或只與其中之一相同。例如,極端嗜鹽細菌能行,但其光合作用色素並非葉綠素類的分子,而是與動物視網膜上的視紫紅質相似的視紫紅質。
原來以為有細胞形態的生物只有原核細胞和真核細胞兩大類。自從發現古細菌以後,才將生物分為上述3大類,這就為探索生命起源和真核細胞起源提供了新的線索。
古菌、細菌和真核生物
在細胞結構和代謝上,古菌在很多方面接近其它原核生物。然而在基因轉錄這兩個分子生物學的中心過程上,它們並不明顯表現出細菌的特徵,反而非常接近真核生物。比如,古菌的轉譯使用真核的啟動和延伸因子,且轉譯過程需要真核生物中的TATA框結合蛋白和TFIIB。
古菌還具有一些其它特徵。與大多數細菌不同,它們只有一層細胞膜而缺少肽聚糖細胞壁。而且,絕大多數細菌和真核生物的細胞膜中的脂類主要由甘油酯組成,而古菌的膜脂由甘油醚構成。這些區別也許是對超高溫環境的適應。古菌鞭毛的成分和形成過程也與細菌不同。
基於rRNA序列的系統發生樹,顯示了可明顯區別的三支:細菌(Bacteria)、古菌(Archaea)和真核生物(Eukarya)
與真細菌主要區別
1、形態學上,古細菌有扁平直角幾何形狀的細胞,而在真細菌中從未見過。
2、中間代謝上,古細菌有獨特的輔酶。如產甲烷菌含有F420,F430和COM及B因數。
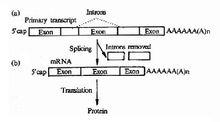
3、有無內含子(introns)上,許多古細菌有內含子。
4、膜結構和成分上,古細菌膜含醚而不是酯,其中甘油以醚鍵連線長鏈碳氫化合物異戊二烯,而不是以酯鍵同脂肪酸相連。
5、呼吸類型上,嚴格厭氧是古細菌的主要呼吸類型。
6、代謝多樣性上,古細菌單純,不似真細菌那樣多樣性。
7、在分子可塑性(molecular plasticity)上,古細菌比真細菌有較多的變化。
8、在進化速率上,古細菌比真細菌緩慢,保留了較原始的特性。
