
簡介
中文名稱:共軛亞油酸
 共軛亞油酸
共軛亞油酸英文別名:10E,Z12-CLA; (10E,12Z)-10,12-OCTADECADIENOIC ACID; 10E,12Z-CLA; 10(E),12(Z)-OCTADECADIENOIC ACID; 10TR-12C-OCTADECADIENOIC ACID; 10-TRANS, 12-CIS CLA; LINOLEIC ACID (10-TRANS, 12-CIS); LinoleicAcid; TRANS-10,TRANS-12-OCTADECADIENOICACID; CIS-10-CIS-12-CONJUGATEDLINOLEICACID; CIS-10,CIS-12-OCTADECADIENOICACID; CLA; Conjugated linoleic acid; (9Z,12Z)-octadeca-9,12-dienoic acid; (2E,4E)-octadeca-2,4-dienoic acid; (9E,11Z)-octadeca-9,11-dienoic acid; octadeca-2,4-dienoic acid; octadeca-10,12-dienoic acid
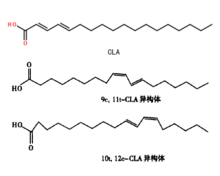 共軛亞油酸分子式
共軛亞油酸分子式CAS:2420-56-6;121250-47-3
EINECS:200-470-9分子式:C18H32O2
主要作用
共軛亞油酸作為一種新發現的營養素,目前在歐美的健康食品界,幾乎已經成了預防現代文明病的萬靈丹,從抗癌到預防心血管疾病、糖尿病,到體重控制上,幾乎是生活在二十一世紀現代人不可或缺的健康食品。
1.共軛亞油酸(CLA)由紅花提煉,是一系列雙鍵亞油酸,具有清除自由基,增強人體的抗氧化能力和免疫能力,促進生長發育,調節血液膽固醇和甘油三酸脂水平, 防止動脈粥樣硬化,促進脂肪氧化分解,促進人體蛋白合成,對人體進行全面的良性調節等作用。
2.共軛亞油酸(CLA)顯著增加人體的心肌肌紅蛋白、骨骼肌肌紅蛋白含量。肌紅蛋白對氧的親和力比血紅蛋白高六倍。由於肌紅蛋白的快速增加,大大提高了人體細胞貯存及轉運氧氣的能力,讓運動訓練更有效,人體活力更充沛。
3.共軛亞油酸(CLA)能增強細胞膜的流動性,防止血管皮質增生,維持器官微循環的正常功能,維持細胞的正常結構及功能,增強血管的舒張能力,有效防止因嚴重缺氧造成的人體臟器和大腦的損傷,尤其是顯著抑制因嚴重缺氧造成的肺、脾水腫。
4.據國外一份專利指出,CLA能有效的發揮“血管清道夫”的作用,可清除血管中的垃圾,有效調節血液黏稠度,達到舒張血管、改善微循環、平穩血壓的作用。有專家還認為,CLA具有擴張和鬆弛血管平滑肌、抑制血液運動中樞的作用,降低了血液循環的外周阻力,使血壓下降,尤其是使舒張壓下降更為明顯。
5、免疫調節功能:CLA會通過以下幾種方式改善免疫相關的反應: 調節腫瘤壞死因子-Α、細胞因子( 白細胞介素1,4,6,8)、前列腺素或氮氧化物,同時減少過敏性免疫反應。
6、改善骨質
減肥功效
近來有越來越多的臨床研究報告都證明,CLA在體重控制上的卓越表現,因為一般人減肥都只會著重在減"重"而非減"肥",也就是說體脂肪比率不一定會有改變,減肥的人如果能配合CLA的使用,可以有效的降低體內脂肪組織相對於瘦肉組織的比率,真正的減到肥肉,而使瘦肉比率上升,這樣的好處是瘦肉越多,體內新陳代謝能力越高,於是形成良性循環,減肥將更容易達到目標。另外,臨床上發現,服用CLA的減肥者,情緒穩定性較高,比較能持之以恆的進行減肥計畫,睡眠和精神狀況也比較好,還有研究報告指出,CLA可以避免減肥者出現溜溜球(yoyo)症狀,使減肥者不至於陷於體重減而復胖,胖而復減的惡性循環中。目前部分減肥食品中已經添加了共軛亞油酸,是一種天然減肥產品。
相關研究
人們早就知道有共軛亞油酸這種物質,但它的重要生理功能的發現,還是上個世紀的後期。大量的科學研究證明,共軛亞油酸具有抗腫瘤、抗氧化、抗動脈粥樣硬化、提高免疫力、提高骨骼密度、防治糖尿病等多種重要生理功能;而且還能降低動物和人體膽固醇以及甘油三酯和低密度脂蛋白膽固醇、還可以降低動物和人體脂肪、增加肌肉。
適宜人群
體重超標者、減脂人群、運動員或運動愛好者
血脂過高的人士、免疫力不佳者
共軛亞油酸在反芻動物中的生物合成與營養控制CLA的獨一無二性與日糧中不飽和脂肪酸在瘤胃中不完全生物加氫有關。微生物對日糧亞油酸的不完全氫化作用共軛亞油酸的一種生成途徑是日糧中不飽和脂肪酸(PUFA)在反芻動物瘤胃中經微生物不完全氫化產生。
Kepler等(1967)發現瘤胃微生物(主要是厭氧的溶纖維丁酸弧菌)可利用多不飽和脂肪酸產生CLA。當日糧脂質被反芻動物消化時,亞油酸在瘤胃中進行兩種重要的轉化作用。第一步:微生物酯酶催化酯鍵水解。這一步是進行第二次轉化—USFA生物加氫的前提條件。第二步反應是Cis-9,trans-11CLA轉變為trans-11C18∶1。與亞油酸生物加氫類似,亞麻酸的生物加氫也是由異構化開始,接著是一系列雙鍵減少,最終生成硬脂酸。
亞油酸和亞麻油酸在瘤胃微生物的氫化作用下,也能發生位置異構化形成CLA(Pariza等,1997),當氫化不完全時,一部分可過瘤胃而直接進入腸道被吸收,因此也稱為“過瘤胃脂肪酸”。c9,t11-CLA異構體就是日糧亞油酸在瘤胃為微生物氫化而產生。所以,在日糧中添加富含CLA或亞油酸的油脂類飼料,可以增加合成CLA的底物量,從而提高CLA的含量。
共軛亞油酸的內源合成共軛亞油酸在反芻動物體內的合成還存在另一條途徑,即經由11-十八烯酸經肝臟和乳腺Δ9脫氫酶脫氫而內源合成,並且CLA在乳腺中的內源合成要比瘤胃微生物氫化作用產生的CLA多。
肝臟微粒體中一種脂類代謝中常見的組織酶—Δ9去飽和脫氫酶,可以使11-十八烯酸去飽和而形成CLA,這是內源合成CLA的重要機制。Santora等(2000)給大鼠飼餵11-十八烯酸,發現51%的CLA沉積到組織中,並經脫飽和作用形成CLA。Kay等(2003)給奶牛皺胃灌注蘋婆酸(Δ9去飽和酶的抑制劑),乳脂中的CLA降低71%,表明乳脂中的大部分CLA系內源合成。
Griinari等(2000)研究發現,CLA可通過體內的Δ9脫氫酶而由內源途徑產生,即由反-11-十八烯酸轉化而來,通過奶牛的真胃瘺管灌注反-11-十八烯酸,結果使奶牛的CLA提高了31%。可見,CLA主要是經由反-11-十八烯酸的內源合成。此外,瘤胃中的一些厭氧菌含有Δ9脫氫酶,能夠將反-11十八烯酸通過Δ9脫氫轉化為c9,t11-CLA。而對於CLA的異構體而言,t10,c12-CLA被認為是瘤胃發酵的結果,尚無存在Δ10脫飽和酶的報導(Baugmard等,2000)。c9,t11-CLA是在乳腺中內源合成的,添加CLA對其無影響,而t10,c12-CLA是由日糧添加CLA合成的。
牛乳脂中CLA的含量反芻動物產品中的CLA含量較高,並且90%以上是具有生物活性的cis-9,trans-11異構體(Parodi,1977和1994)。乳中的CLA的含量相差很大,占脂肪含量的0.3%~0.6%左右(Dhiman等,2000)。目前人們通過飲用牛奶獲取的CLA的量遠低於經動物實驗證明具有特殊生理功能的人體需要量,因此,如何獲取更多的CLA以促進人們的身體健康成為科學家極為關注的課題。
通常,增加CLA攝入量的方法有兩種,一是通過改變飼料配方和飼養方法,提高畜產品中的CLA的含量。另一種方法是在食品中,尤其是在乳製品或者肉製品中,進行CLA的強化,提高食品中CLA的含量。影響乳脂中共軛亞油酸含量的日糧因素乳脂CLA主要來源於18碳PUFA瘤胃氫化的中間產物cis-9,trans-11C18∶2和trans-11C18∶1,凡是有利於使這兩種中間產物到達真胃的量增加的因素均可以促進乳脂CLA含量的提高。
如上所述,反芻動物源性食品中脂肪的共軛亞油酸含量取決於瘤胃中共軛亞油酸與trans11,C=18∶1的產量和組織中Δ9脫氫酶的活力。許多日糧因子影響乳脂中共軛亞油酸含量,根據作用的潛在機理可分為三類:第一類是向瘤胃提供生成共軛亞油酸或trans11,C18∶1脂質底物;第二類是能夠改變瘤胃內環境,進而影響與瘤胃生物加氫有關的細菌;第三類既能提供脂質底物,又能改變瘤胃菌群。
脂質底物在一定添加範圍內,乳脂CLA含量與油脂添加量成正相關。Dhiman等(2000)發現,豆油添加量在乾物質進食量的4%範圍內時,奶牛乳脂CLA含量隨豆油添加量的增加逐漸上升;Mir等(1999)在山羊中所做的研究顯示,乳脂CLA含量隨菜子油添加量增加而逐漸提高。不同油脂的添加效果不同,這主要決定於脂肪酸的組成。Kelly等(1998)給奶牛補飼5.3%的花生油、葵花子油和亞麻子油分別使乳脂CLA濃度達到每克13.3毫克、24.4毫克和16.7毫克。花生油和葵花子油分別含有不足30%和超過60%的亞油酸,亞麻酸都幾乎為0,在相同添加水平下對乳脂CLA含量的提高效果差異基本反應了這一組成差異。亞麻子油的亞油酸和亞麻酸總含量與葵花子油的亞油酸含量相差不大,可對乳脂CLA含量的改善效果差異顯著,這顯示亞麻酸提高乳脂CLA的效率不如亞油酸。
反芻動物日糧中添加植物油對瘤胃細菌生長有抑制作用,飼餵脂肪酸鈣可把這種抑制作用降低到最低限度。飼餵全脂子實也可以降低這種抑制作用,但由於瘤胃細菌幾乎不能利用未加工子實中的多不飽和脂肪酸,因而飼餵完整子實對乳脂中共軛亞油酸含量沒有作用。若全脂子實進行適當加工,乳脂共軛亞油酸含量將會大量增加,例如將油菜子、大豆、棉子進行破碎、烘烤和擠壓加工等。Dhiman等(1999)給奶牛補飼全脂膨化大豆和全脂膨化棉子,分別使CLA含量由每克3.6毫克提高8.6毫克和7.2毫克,補飼高油玉米對乳脂CLA沒有影響(Dhi鄄man等,1999)。補飼18%的破碎烘烤大豆和3.6%豆油後,奶牛日糧脂肪含量和脂肪中亞油酸與亞麻酸組成很接近,但乳脂肪酸中CLA的濃度差別很大,分別為7.7和21.0mg/g(Dhiman等,2000)。
Chouinard等(1998)給奶牛分別補飼17.5%的粉碎生大豆、膨化大豆、微粉化大豆和烘烤大豆,相應的乳脂CLA含量分別為每克3.0毫克、8.9毫克、7.0毫克和6.6毫克。Dhiman等(2000)補飼破碎生大豆沒有對乳脂CLA產生影響,而補飼破碎烘烤大豆使乳脂CLA提高了1倍。不同加工處理方式可能改變了大豆脂肪酸在瘤胃釋放的速率和比例(Dhiman等,2000)。
瘤胃環境調控劑補飼魚油也可使牛乳脂的CLA含量提高。給奶牛餵以乾物質計0%、1%、2%和3%的魚油,導致採食量、產奶量、乳脂率下降,但增加了乳中CLA含量。餵3%魚油奶牛乳脂肪每克含22.5毫克CLA脂肪(Donovan等,2000)。餵3%(以乾物質計)魚粉與無魚粉飼料比較,奶牛乳脂肪中CLA含量增加63%(Dhiman等,1999)。Scollan等(1997)報導補飼魚油或魚粉會導致trans-11C18∶1
在瘤胃聚集。魚的種類繁多,魚油的脂肪酸組成隨魚品種的不同而有一些差異,但從部分魚油的脂肪酸組成看,各種魚油在脂肪酸組成上具有一些共性。魚油的18碳PUFA並不豐富,尤其是亞油酸和亞麻酸,總含量不超過5%,不過魚油中還含有少量C18∶4,油酸含量在10%~20%之間,20碳以上的UFA含量豐富,在27%~48%之間。由於魚油的亞油酸和亞麻酸含量只比動物脂略豐富一點,因此瘤胃聚集的trans-11C18∶1究竟來自哪裡尚不清楚。有學者認為,可能魚油對瘤胃中反十八碳烯酸生物加氫的抑制作用與前述高水平亞油酸的抑制作用類似。也有學者認為,飼餵魚油導致瘤胃反十八碳烯酸可能是由於抑制了減少反十八碳烯酸細菌的生長或專一性地抑制了這些細菌產生酶的作用。(見表1)瘤胃菌群的改變常常是瘤胃pH值降低的結果,以丙酸產量的增加為標誌。高精低粗日糧可提高瘤胃及乳脂中反十八碳烯酸的含量,向低粗料日糧中添加緩衝劑可提高瘤胃pH值,從而降低反十八碳烯酸的產量。離子載體可抑制溶纖維丁酸弧菌等革蘭氏陽性菌的生長,從而抑制亞油酸的生物加氫,導致瘤胃內容物硬脂酸減少而C18∶1增加。但是,奶牛日糧中添加離子載體不一定能增加乳脂中的共軛亞油酸含量,這可能與瘤胃細菌的適應能力有關。
混合因素與飼餵含類似脂質的全混合日糧相比,牧草可提高乳脂的CLA含量。
在補飼富含亞油酸的脂肪飼料的同時添加離子載體,可以降低瘤胃trans-11C18∶1與CLA的比值(Fellner等,1997)。在其研究中,灌注亞油酸的同時添加離子載體使trans-11C18∶1/CLA下降一半以上,CLA在總脂肪酸中的比例則提高了2倍~3倍。考察的三種離子載體中,莫能菌素的影響最小。
Dhiman等(1999)發現莫能菌素處理對乳脂CLA含量的影響不顯著,不過有提高CLA的趨勢。Chouinard等(1998)也沒有發現添加莫能菌素對乳脂CLA含量的顯著作用。
Kelly等(1998)發現全部營養來自優質牧草的奶牛乳脂CLA含量是配合飼料組奶牛的兩倍。Dhiman等(1999)的研究顯示,對照組(以玉米和玉米青貯料為飼料)、1/3、2/3或全部飼料為新鮮牧草的試驗組奶牛的乳脂CLA含量分別為每克3.8毫克、8.9毫克、14.3毫克和22.1毫克乳脂肪酸。Jahreis等(1997)比較了不同飼養制度對CLA含量的影響,舍飼組、傳統牧場放牧組和生態牧場放牧組的CLA含量分別為每克3毫克、6毫克和8毫克乳脂肪酸,顯然造成CLA含量差異的根源在於新鮮牧草採食量及其質量。
以乾物質為基礎,青綠牧草的脂肪酸含量在1%~3%之間,其中90%以上為UFA。表2列出了一些粗飼料的18碳UFA組成,不同品種牧草的組成有一定的差異。總體來看,新鮮牧草的C18∶4含量豐富,占一半以上,其次是C18∶3。牧草中所含C18∶2的雙鍵分別在第7和第9位,不具有通過瘤胃氫化產生乳脂CLA前體的功能,因此新鮮牧草對乳脂CLA含量的促進作用可能主要來自於C18∶4和C18∶3。從UFA組成看,乾草C18∶4和C18∶3含量也很豐富,但與青綠飼料相比,飼餵乾草時乳脂CLA含量明顯較低,這可能是由於乾草的總脂肪酸含量較低造成的,許多乾草的粗脂肪含量不足1%。牧草提高乳脂的CLA含量的機理可能是脂質底物與牧草組分的協同作用改變了瘤胃內的生物加氫過程所致。
總結由於CLA對許多人類頑固疾病具有預防和治療作用,因此,如何將其作為一種功能性食品的開發也日益受到人們的關注。但相關的研究表明,利用人工方法對亞油酸或富含亞油酸的植物油進行催化而合成CLA不僅費用較高,而且合成的CLA仍然是多種異構體的混合物,大量分離純化得到單個異構體的技術難題尚未解決。
因此較為簡單、經濟的方法是從動物產品中獲得CLA。反芻動物產品尤其是牛乳中含有豐富的CLA,並且主要是具有生物活性的c-9,t-11異構體。因此,研究CLA與乳脂合成代謝的關係,探索通過飼料營養措施生產CLA功能牛奶,不僅能提高牛奶營養學價值.
改善牛奶品質,而且也能為人類健康保健提供新的途徑和產品。
物質來源
共軛亞油酸(CLA)是一種主要從反芻動物脂肪和牛奶產品中發現的天然活性物質,是一類含有共軛雙鍵的十八碳二烯酸(亞油酸)異構體混合物。亞油酸和亞麻酸在反芻動物瘤胃內通過異構化和生物脫氫反應形成CLA,反式脂肪酸在動物細胞內經△9-脫氫酶的脫氫作用也能形成CLA。飼料、瘤胃微生物、瘤胃pH值以及品種等都對CLA有著重要的影響。反芻動物來源的食品是共軛亞油酸最主要的天然來源。
