簡述
1986年APC基因首次由Herrera在一位患有直腸腫瘤及智力缺陷的Gardner綜合徵的患者染色體上發現,該患者的5號染色體長臂上有一段缺失. 隨後發現APC基因是FAP的致病基因. APC基因是一個很大的管家基因(housekeeping gene),全長含有一個8538 bp的開放式可讀框架,共15個外顯子,6個可變地表達. 其中第15外顯子獨自含有6571個bp,組成77%的編碼區,是人類已知最大的外顯子,它共編碼2843個胺基酸. 轉錄產物mRNA分子為8.9 kb,在很多細胞和組織中均有表達. Miyoshi等根據在多種腫瘤(包括結腸直腸腫瘤)中常見雜合子5q21的缺失,將APC基因歸為腫瘤抑制基因. 在微細胞融合技術中,通過向結腸癌細胞系中轉染含5q21區的正常第5號染色體,可逆轉該腫瘤的惡性表現. 然而一旦去除轉染的第5號染色體,該細胞系又恢復了惡性行為,這表明5q21區確實存在著結腸癌的抑制基因. 另一方面,在FAP癌變患者中,5q21區高頻發生的雜合性丟失(loss of heterozygosity, LOH)現象也支持了APC基因屬於腫瘤抑制基因.
基因的功能
APC 蛋白與 β連環蛋白 ( beta-catenin ) ( 一種轉錄transcription 因子 ) 形成複合物,導致β連環蛋白降解。如果缺乏 APC 蛋白,過多的β連環蛋白就會在 細胞核 (nucleus) 內的聚集。β連環蛋白與細胞核內的另一種蛋白結合 , 形成一種複合物;這種複合物又與 DNA結合,啟動了幾種 基因 (genes) 的轉錄。這種複合物中的一個靶基因叫做c-myc一種已知的原癌基因。C-myc本身就是幾種基因的轉錄因子 (transcription factor), 它控制著細胞的生長和分裂。因此,APC基因的突變導致了一系列的連鎖反應,最終導致細胞分裂的加速。
研究顯示, 對缺乏APC 蛋白的結腸癌細胞添加APC 蛋白質有減退腫瘤細胞成長功能。成長的減退是由於細胞自動死亡增加的結果, 可見 APC 有控制細胞死亡並且成長的功能. 所以, APC基因的損失對細胞成長與細胞死亡之間的平衡有一定的影響。 APC基因控制細胞數量。
基因突變
![APC[腫瘤抑制基因APC]](/img/9/14e/nBnauM3X2MDOwcDO3cDO4UjM2UTM1QDN5MjM5ADMwAjMwUzL3gzLwAzLt92YucmbvRWdo5Cd0FmLyE2LvoDc0RHa.jpg) APC[腫瘤抑制基因APC]
APC[腫瘤抑制基因APC]APC基因突變類型主要有點突變和框架移碼突變,前者包括無義突變,錯位突變和拼接錯誤,後者包括缺失和插入. APC基因突變有300多種,這些突變遍及整個基因,60%以上的突變位於第15號外顯子的5′端. 其中密碼子第1286~1513號之間的10%左右的編碼區集中了約65%的體細胞突變,被稱為突變密集區(MCR). 大部分突變屬於錯位突變,由缺失或1~8個鹼基對的插入引起. 大約95%的突變結果是在下游形成提前的終止密碼子,使APC蛋白呈截短改變,這可能削弱了APC蛋白固有的抑制細胞增殖功能,從而導致APC蛋白功能的障礙;Jarvinena等[1]認為這些APC截短蛋白可以結合於野生型APC蛋白,並對野生型蛋白具有顯性負效應, 使之失活而導致腫瘤發生. 現國內外關於APC基因突變的研究很多,不同的報導所選擇的研究目的基因也大都不相同,但大部分以研究15號外顯子突變為主. Miyaki等[2]報導日本人在MCR以密碼子1260~1453段突變率最高;國內王艷等[3]報導在1339~1436密碼子區域突變率最高,1260~1359密碼子區域突變率最低;而高楓等[4]將1337~1453段分為兩段,分別設計引物擴增,也證實了上述說法.
APC蛋白
APC基因編碼蛋白複合物可分成兩個大區:羧基端區占75%,氨基端區占25%. 靠近氨基端的部分含有大量亮氨酸殘基並具有與肌球蛋白、中間絲蛋白局部同源的序列;靠近羧基端的區域含有較多的絲氨酸殘基、酸性胺基酸和脯氨酸殘基. 這兩個區域記憶體在螺旋螺旋(coiledcoils)結構,提示蛋白與蛋白之間的相互作用. APC蛋白的結合特性的研究提示,氨基端是同質寡聚反應的關鍵. 另外,起始的171個殘基對複合物形成起主要作用,在這171個殘基中,前45個殘基又是關鍵. 大多數APC基因突變產生的蛋白為171殘基之外的剪下所致. 一般說來,截去末端的蛋白仍保留繼續寡聚的潛力,還可構成滅活複合物,這就從本質上說明了突變蛋白的優勢效應. APC蛋白的主要作用是與β連環蛋白(βcatenin)和E鈣黏附蛋白相互作用而影響細胞黏附及細胞間信號傳遞,是β連環蛋白的負性調節子. β連環蛋白基因位於3q21,編碼蛋白分子質量約92 ku. 蛋白結構包含有α連環蛋白、APC和E鈣黏附蛋白的結合位點. 高水平的β連環蛋白可通過GSK3β使得APC蛋白磷酸化,從而促進其對β連環蛋白的降解效率,使胞質內β連環蛋白水平保持在一種平衡狀態. 通過對APC基因突變的研究,人們發現,部分APC基因MCR的突變,可導致APC蛋白雖能與β連環蛋白結合,但卻不能降解β連環蛋白[5],提示在腫瘤發生中,APC基因突變的一個關鍵意義是失去對β連環蛋白的調節. 因為APC蛋白的β連環蛋白結合位點是高度保守的,證明突變型APC蛋白與β連環蛋白形成複合物的能力在腫瘤的發生中非常重要[6]. 此外,APC蛋白還結合微管,在細胞分裂和移動中起作用. APC可通過調控周期素依賴周期素激酶複合物的活性而調節細胞周期,它還通過誘導凋亡而介導其在結腸腺瘤發生中的作用,故被譽為結腸上皮完整性的分子性“門衛”(molecular "gatekeeper"). APC基因的突變可改變APC蛋白與β連環蛋白及E鈣黏附蛋白之間的平衡,導致細胞細胞、細胞基質之間黏附作用以及接觸抑制信號傳遞的改變,引起細胞分裂與細胞死亡之間的平衡失調,以致生長失控,成為結直腸癌的一個限速的分子因素。
基因突變
APC基因常見於家族性腺瘤性息肉病(FAP). 人類體細胞染色體為2倍體. FAP患者遺傳一條突變APC基因,保留一條正常的等位基因. 不同於典型的孟德爾遺傳疾病,只要保留的正常APC等位基因發生突變,則出現多發性息肉. 有79% FAP患者保留的正常APC等位基因發生突變[8]. Nagase等發現約80%家族有APC基因突變. 對其中176例突變進行分析,發現98%以上的突變可導致APC蛋白截短,這些突變分別表現為為無義突變(33%)、小插入(6%)或缺失(55%). Miyoshi等[9]分別使用限制性片段長度多態性(RFLP)法及聚合酶鏈反應單鏈構象多態分析法(PCRSSCP),在檢測FAP患者中APC基因突變與病理類型之間關係時發現,雖然FAP輕中度腺瘤存在著APC基因的胚系突變,但APC基因的LOH發生率卻極低. 然而自重度腺瘤向黏膜內癌侵潤發展的各病理類型中,LOH的發生率顯著增加. 而且在侵潤癌中,觀察到了APC基因胚系突變與LOH同存的現象. 這些現象表明了APC突變基因的雜合子狀態足以使FAP早中期腺瘤的形成,而APC基因突變加上雜合性丟失,則與向癌的進一步轉化有關.
此外,突變定位與FAP的臨床表現也有一定聯繫. 研究發現,APC基因最靠近5′端的突變(在外顯子4或更近側)產生輕型腫瘤表型,而靠近3′端MCR的突變產生重型表型[10].帶狀瘤與15號外顯子的1445~1578密碼子之間的突變有關,其近側的突變先天性視網膜色素上皮增生(CHRPE)為陰性,而遠側的突變(在密碼子463~1387之間)CHRPE為陽性;在密碼子1403~1578之間的截短型突變與Gardner綜合徵有關;而另外一種縮減型息肉病(attenuated FAP,AFAP)患者的突變常位於APC基因第3,4號外顯子的5′末端和密碼子1578的3′端[11].基因型與表型之間的關聯可以用正常APC分子間的二聚體作用來解釋. 遠側的突變靠近3′端,產生一段較少截短的蛋白質,可與野生型APC蛋白質結合併干擾其功能(即顯性負效應);而靠近5′端的突變產生嚴重截短的蛋白質,不能二聚體化,使野生型蛋白質保持正常功能. 除該病表現的差異外,息肉數目也與突變位點有一定聯繫. 通常認為,息肉數目1000以下為稀疏型,數目在5000以上為密集型.密碼子1249附近的突變及密碼子1465遠端突變與稀疏型有關,而密碼子1250與1330之間的突變與密集型有關. 此外,密碼子1309點突變與胃腸道症狀較早出現有關.
雖然APC基因突變直接導致FAP發生,但其意義不只限於FAP病,在非FAP的散發性結直腸腫瘤中同樣觀察到了APC基因的突變[12]. 這表明APC基因突變不僅與FAP發生有關,而且涉及非FAP的散發性結直腸腫瘤. 並且,APC基因突變發生於小於1 cm腺瘤的事實,提示APC基因突變是結直腸腫瘤發生過程中的早期事件.
檢測方法
通過檢測APC基因突變,可篩選出FAP家族成員高危患者,也可用於評估結直腸癌的療效和預後,所以檢測APC基因變化對預防、早期診斷及早期治療結直腸癌具有重大意義[13]. 檢測APC基因突變的方法報導較多,大致分為3類.
第1類,直接序列分析,PCRSSCP法. 檢測APC基因高度多態性DNA標記,不必對整個龐大的APC基因所有外顯子進行檢測[14]. 套用微衛星多態性標記進行LOH分析是目前抑癌基因定位和等位基因丟失研究中最經常採用的方法. APC基因等位丟失亦常出現在散發性結直腸癌中,LOH發生率為35%~45%[15]. 此類方法適合於較小片段基因(100~500 bp)突變的檢測,對APC基因需分成40個相互重疊的片段來檢測,費時,費力.
第2類,包括異源脫氧核糖核酸分子(DNA)分析法(Hdxa)和錯配化學清除/羥胺鋨酸酐(CCM/HOT)等. 一次可檢測較大片段的DNA,減少受檢片段,如CCM/HOT法使之降到25個片段. 然而,仍要對外顯子進行突變檢測,需較大片段的序列分析來確定,對APC基因檢測也非簡便的方法.
第3類,是針對基因突變產生終止密碼而表達不全蛋白質(截短蛋白質)來設計的. 如體外合成蛋白質試驗(IVSP法)和藍/白選擇試驗,可以檢測大片段的DNA或RNA,又能選擇性發現產生終止密碼的突變,與藍/白選擇試驗不同的是,IVSP法還可根據不全蛋白大小來確定突變的位點. 通過對APC基因進行體外合成蛋白質及等位基因特異性表達的測定,可以更有效地檢測到不能被PCRSSCP法所檢測到的APC基因突變.
總之,由於APC基因突變與FAP早期發生密切相關,且FAP具有較高的遺傳傾向,這種疾病如果不加治療,其惡變率幾乎達到100%,所以對APC基因的定位及其功能的研究對於FAP的早期診斷和治療顯得尤為迫切和重要[16]. 如果說最初APC基因的發現為FAP的分子診斷學提供了理論基礎,現APC基因的研究進展正使針對這種疾病的分子診斷方法成為可能. 相信通過對APC基因的進一步研究,不僅會有更實用的基因檢測診斷方法問世,而且可能得到針對腫瘤發生機制的特異性基因治療方法
APC啟動子有兩個區,1A和1B。啟動子1A區最為活躍。APC啟動子高甲基化也可能是導致結直腸癌的原因。有研究表明APC啟動子1A區在結直腸癌中高甲基化,二在腺瘤中不甲基化。APC啟動子高甲基化抑制了APC的轉錄。據報導APC的1A啟動子區高甲基化也發生在胃腸癌症中,比如食管癌,胃癌,胰腺癌,肝癌,結直腸癌。
抑癌基因
| 抑癌基因 | 功能 | 相關腫瘤 |
| Rb | 轉錄調節因子 | RB、成骨肉瘤、胃癌、SCLC、乳癌、結腸癌 |
| p53 | 轉錄調節因子 | 星狀細胞瘤、膠質母細胞瘤、結腸癌、乳癌、成骨肉瘤、SCLC、胃癌、磷狀細胞肺癌 |
| WT | 負調控轉錄因子 | WT、橫紋肌肉瘤、肺癌、膀胱癌、乳癌、肝母細胞瘤 |
| NF-1 | GAP,ras GTP酶激活因子 | 神經纖維瘤、嗜鉻細胞瘤、雪旺氏細胞瘤、神經纖維瘤 |
| DCC | 細胞粘附分子 | 直腸癌、胃癌 |
| p21 | CDK抑制因子 | 前列腺癌 |
| p15 | CDK4、CDK6抑制因子 | 成膠質細胞瘤 |
| BRCA1 | DNA修復因子,與RAD51作用 | 乳腺癌、卵巢癌 |
| BRCA2 | DNA修復因子,與RAD51作用 | 乳腺癌、胰腺癌 |
| PTEN | 磷酯酶 | 成膠質細胞瘤 |
| APC | WNT信號轉導組分 | 結腸腺瘤性息肉,結/直腸癌 |
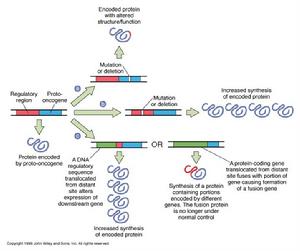 原癌基因激活的可能途徑
原癌基因激活的可能途徑