![雙元系統[信號轉導系統]](/img/7/b44/nBnauM3X4MTO2gDMyQTMzUTN1UTM1QDN5MjM5ADMwAjMwUzL0EzL1EzLt92YucmbvRWdo5Cd0FmLzE2LvoDc0RHa.jpg "雙元系統[信號轉導系統]")
受體組成
組氨酸激酶(HK)
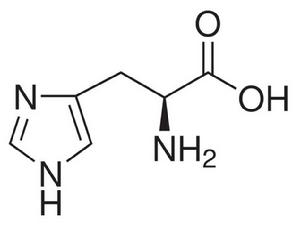 組氨酸(His)結構式
組氨酸(His)結構式組氨酸蛋白激酶(histidine kinase,HK)位於質膜,分為感受胞外刺激的 信號輸入區域和具有激酶性質的 轉運區域。當輸入區域接受信號後,轉運區域激酶的組氨酸殘基(His)發生磷酸化,並將磷酸基團傳遞給下游組分。
細菌組氨酸蛋白激酶的催化結構域(catalytic domains)不同於絲氨酸、蘇氨酸或酪氨酸激酶,而與n型拓撲異構酶旋轉酶B(type I topoisomerase gyrase B)和伴侶蛋白Hsp90(chaperone Hsp90)的ATP酶結構域(ATPase domains)相似。推測這是蛋白質中高度保守的谷氨酸殘基參與ATP酶的
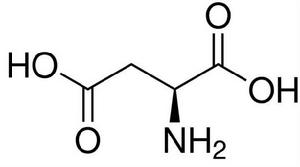 天冬氨酸(Asp)結構式
天冬氨酸(Asp)結構式催化機制,而在組氨酸激酶活性中心沒有發現谷氨酸。以上結構特點可以解釋為什麼這個超家族的成員中有些有激酶的功能,而有些有ATP酶的功能。HPK的主要功能是催化ATP依賴的自磷酸化反應,使二聚體結構域特異的His殘基磷酸化,進而作為磷酸供體使RR蛋白的Asp殘基磷酸化。
應答調控蛋白(RR)
應答調控蛋白(response-regulator protein,RR)也有兩個部分:一個是 接收區域,由天冬氨酸殘基(Asp)接受磷酸基團;另一部分為 信號輸出區域,將信號輸出給下游組分(通常是轉錄因子),以此調控基因表達。
系統結構
細菌
細菌為原核生物,沒有核膜,故其雙元系統介導的信號通路也較為簡單。
| 感應蛋白 (組氨酸激酶)HK | 輸入區域 |
| ↓ | |
| 轉運區域(His) | |
| ↓ | |
| 應答調控蛋白 RR | 接收區域(Asp) |
| ↓ | |
| 輸出區域 |
植物
 植物細胞跨膜信號轉導還需經過核膜
植物細胞跨膜信號轉導還需經過核膜植物細胞為真核細胞,與原核細胞相比,信號通路更加複雜,在HK和RR之間會增加一個或多個傳遞磷酸基團的組分。主要區別有兩點:
HK轉運區域下游增加了一個接收區域,具有可以傳遞磷酸基團的天冬氨酸殘基;
在RR上游增加了一個組氨酸磷酸轉移蛋白(Hpt),它接收HK傳來的磷酸基團後,進一步傳遞給下游的RR。
1.HK轉運區域下游增加了一個接收區域,具有可以傳遞磷酸基團的天冬氨酸殘基;
2.在RR上游增加了一個組氨酸磷酸轉移蛋白(Hpt),它接收HK傳來的磷酸基團後,進一步傳遞給下游的RR。
這樣,複雜的雙元系統實質上是增加了傳遞磷酸基團的蛋白組分。
| 雜合感應蛋白 (組氨酸激酶)HK | 輸入區域 |
| ↓ | |
| 轉運區域(His) | |
| ↓ | |
| 接收區域(Asp) | |
| ↓ | |
| 組氨酸磷酸轉移蛋白(Hpt) | Hpt區域(His) |
| ↓ | |
| 應答調控蛋白 RR | 接受區域(Asp) |
| ↓ | |
| 輸出區域 |
信號轉導途徑實例
細胞分裂素(CTK)
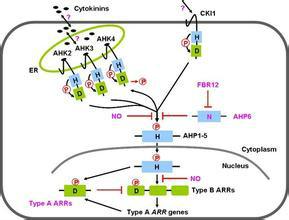 細胞分裂素信號轉導
細胞分裂素信號轉導細胞分裂素受體1( CRE1) 是組蛋白激酶(HPK)類似蛋白。HPK的主要功能是催化ATP依賴的自磷酸化反應,使二聚體結構域特異的His殘基磷酸化,進而作為磷酸供體使RR蛋白的Asp殘基磷酸化。
信號轉導過程中,CTK先於CRE1的HPK部分結合,實現跨膜信號轉換。由CRE1的接收區域D將磷酸基團傳給 組氨酸磷酸轉移蛋白( AHP),AHP進入細胞核後,通過 反應調節蛋白( ARR)引起基因表達,或通過其他效應物引起CTK誘導的生理反應,例如細胞分裂。
乙烯(ethylene)
乙烯受體1( ETR1)定位於內質網,符合乙烯的疏水性,能自由越過質膜進入細胞內。
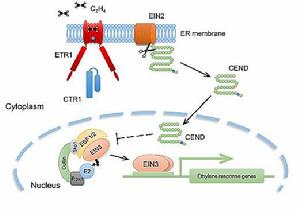 乙烯受體定位於內質網
乙烯受體定位於內質網 水稻乙烯反應及信號轉導
水稻乙烯反應及信號轉導擬南芥中從乙烯受體到細胞核的信號轉導途徑已經初步確定。乙烯與ETR1結合以後,激活了下游的 CTR1。CTR1通過級聯反應將信號傳遞到 EIN2(乙烯不敏感2,ethylene insensitive 2)基因,繼而引發下游的一系列反應。
意義
在動物細胞中,跨膜信號的轉導主要靠G蛋白偶聯受體介導的信號通路來完成。G蛋白是三聚體GTP結合調節蛋白(trimeric GTP-binding regulatory protein)的簡稱,位於質膜內胞質一側,由G、G、G三個亞基組成。其中,G亞基本身具有GTPase的活性,為分子開關蛋白。
然而在植物細胞中,G蛋白偶聯受體介導的信號通路並不常見,雙元系統和受體激酶介導的信號通路才起著更為主要的作用。
