群落穩定性的含義
群落的穩定性具有兩層含義:群落的抗干擾能力,即抵抗力穩定性;群落受到干擾後恢復到原平衡態的能力,即恢復力穩定性。對於群落的穩定性,有平衡說(equilibrium)和非平衡說(non-equilibrium)兩種對立的觀點。前者認為,共同生活在同一群落的物種處於一種穩定狀態,通過競爭、捕食和互利共生等種間相互作用而形成相互牽制的整體,導致生物群落具有全局穩定性特點;在穩定狀態下物種組成和各種群數量都變化不大;群落實際上出現變化是由於環境的變化即干擾所引起。後者認為,組成群落的物種始終處在不斷的變化之中,自然界中的群落不存在全局穩定性,有的只是群落的抵抗性(群落抵抗外界干擾的能力)和恢復性(群落在受干擾後恢復到原來狀態的能力),其重要依據就是中度干擾理論,即中等程度的干擾水平能維持髙的物種多樣性;干擾過於頻繁,則先鋒種不能發展到演替中期,多樣性較低;干擾間隔期過長,演替發展到頂極,多樣性又降低; 只有中等程度的干擾使多樣性維持最高水平,它允許更多的物種入侵和定居。
群落的多樣性是群落穩定性的一個重要尺度。當一個群落有很多物種,而且每個種的個體比較均勻地分布時,物種之間就形成了比較複雜的相互關係。這樣群落對於環境的變化或來自群落內部種群的波動,由於有一個較強大的反饋系統,從而得到較大的緩衝。另外,多樣性高的群落,食物鏈和食物網更加趨於複雜,群落內部的能量流動的途徑更多一些,如果某一條途徑受到干擾堵塞不通時,可能有其他路線予以補償。因此,多樣性髙的群落相對比較穩定。反之,若物種數目少,各個種的相對豐度又不均勻,則群落的多樣性就較低,其穩定性也相對較差。
群落多樣性與穩定性的關係
物種多樣性是指地球上生物有機體(生命)的多樣化程度,可以分為遺傳多樣性、物種多樣性和生態系統多樣性3個層次。對群落而言,物種多樣性尤為重要。物種多樣性有兩種含義:其一是種的數目或豐富度,即一個群落或生境中物種數目的多寡;其二是種的均勻度,即一個群落或生境中全部物種個體數目的分配狀況,它反映的是物種個體數目分配的均勻程度。自20世紀50年代以來群落多樣性和穩定性的關係一直是生態學界爭論的焦點問題,目 前存在兩種不同的理論:MacArthur-Elton假說(群落多樣性和穩定性正相關)和May假說(群落多樣性和穩定性無關)。
MacAnhur-Elton假說的核心是物種較多的群落就可能保持穩定。Mac- Anhur(1955)首次提出了這一觀點,認為自然群落的穩定性取決於兩個方面的因素:一是物種的多少,二是物種間相互作用的大小,而物種的多少對穩定性的。這個觀點可以在物種間的捕食關係中得到實證:當某一物種 數量極多時,必須有大最的捕食者來分散過多的能量,同時又必須有大量被捕食者物種以保證一個物種的數量不會下降過多;當某一物種數量較少時,該物種的每一捕食者必須有大量的替代被食者,以減少壓力。Elton根據物種入侵的研究得出了類似的結論,認為一個相對簡單的植物或動物群落易於受到毀滅性干擾,因而抵禦外來種入侵的能力較弱。
20世紀70年代,MacArthur-Elton假說的生物多樣性越高,穩定性越強的觀點受到了挑戰。May(1973,1976)等認為,隨機構造的複雜系統只有在某一臨界值內穩定,超過該臨界值,系統就不會穩定;在多樣化的系統中,一個生態關係複雜的網路,可導致種群急劇波動,而不是更加穩定;共棲的多物種群落,某物種的波動往往會牽連到整個群落;從減少害蟲種群暴發的意義上講,多樣性與穩定性之間並無一般的相關關係。他們提出了多樣性的產生是由於自然的擾動和演化兩者聯繫的結果,環境的多變性和不可測性使物種產生了繁殖與生活型的多樣化。
Pimm(1984)指出,形成多樣性-穩定性的兩種不同觀點的主要原因在於多樣性、複雜性和穩定性有許多不同的定義和概念。MacArthur-Elton的理論雖已被許多人加以論證,但還有待於深入和提高;May所提出的理論又過多地依賴於數學方法,而且他所使用的系統又是以人工模擬為主,因而離實際相差甚遠。Pimm對多樣性-穩定性關係做了一個總結,認為理論上對於一個有更多物種的群落:要使群落更加穩定,就需要物種間的聯結變得更少;群落內種群的彈性變得更小;當一個物種丟失以後,群落內種類成分和生物量有較大變化;一個物種丟失以後的狀態將保持更長的時間。由此可見,傳統的靜態的內部自調穩定性概念,難以客觀地反映生態系統的動態特徵。 Diamond和Case(1986)提出,生態學本身就是一門“多元學科”,在理論和 方法上採用多元論的觀點是很必要的。現代生態學已由強調群落或生態系統的平衡、穩定性、均勻性、均質性、確定性、單尺度和小尺度現象,發展到開始重視非平衡、不穩定性、異質性、不確定性、多尺度和等級特點,這將有助於更好地理解多樣性-穩定性關係。
群落穩定性的分析方法
食物網與穩定性
MacArthur (1955)根據群落的複雜性與群落的穩定性有關的觀點,認為一個群落的物種越多或食物網的環節越多,群落內部物種之間的能流途徑也就越多,從而群落將更加穩定。於是,他通過一個假想的捕食者與被食者(或害蟲與天敵)的營養聯繫對群落的穩定性作了定量研究。
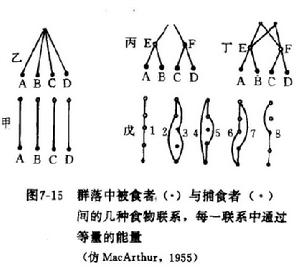 不同群落營養結構
不同群落營養結構圖中甲、乙、丙、丁是四個不同的群落營養結構,其中捕食者與被食者的關係很不一樣。每圖中A、B、C、D代表被食者,E、F、G、H代表捕食者,每一條線表示捕食聯繫, 也就是能流途徑。甲圖中捕食者與被食者的關係十分專一,是單種聯繫;乙圖中捕食者G可以捕食被食者A、B、C、D中的任何一種;丙圖是兩個營養階層的捕食關係,G可以捕食 E、F,而E、F又分別可以捕食A、B和C丁圖是一個錯綜複雜的二個營養階層交錯的捕食關係。假設每條食物鏈能流的量都相同,則甲、乙、丙的食物鏈都是四條,甲為 AE、BF、CG 和 DH,乙為 AG、BG、CG 和 DG,丙為 AEG、BEG、CFG 和 DFG,每 一條能流的量各占1/4;而丁的食物鏈有八條,即 AEG、AEH、BEG、BEH、CFG、CFH、 DFG和DFH,每一條能流的量為1/8。
MacArthur的理論和方法,雖然得到不少學者的贊同和支持,如Paine (1966, 1969) 提供的材料。但是,這個方法目 前在實際工作中套用很少,理論上也受到挑戰,因為能流在食物網中的分配是不均勻的,難以準確地測定每一條食物鏈所占有的能量比例。
群落動態與穩定性
70年代以來,由於新興橫斷學科的發展和不同學科之間的相互滲透,不少學者(如 Angelis,1975; May,1971,1976; Svirezhevetal,1978)基於描述群落動態特性的數學模型,採用系統理論中的Mapunov穩定性定理來分析和探討生物群落的穩定注。
