穩態轉化概念是從人們對自然生態系統存在多暫態平衡的直觀認識中抽象出來的。一般來說,自然界的生態系統都會具有兩個或更多的可交替(或可轉化)的穩定狀態。在淺水湖泊中,水生高等植物占優勢的清水狀態和浮游藻類占優勢的濁水狀態便是生態系統多穩態的很好的例子。淺水湖泊中二種穩定狀態(清水穩態 vs 濁水穩態)之間的相互轉化,基於三個經驗事實:1)湖水的濁度隨營養鹽水平的增加而增加,2)沉水植被降低濁度,3)當濁度超過某一臨界點時,沉水植被消失。濁度和營養水平之間存在二種不同的函式關係,一種是水生植被占優勢的情形,另一種是無植被的情形。在較低的營養鹽水平下,只有植被占優勢的平衡存在,而在較高的營養鹽水平,僅有無植被的平衡存在。在一個中間的營養鹽範圍內,二種平衡都存在:一種有植被,而另一種較混濁而無植被,它們被一個不穩定的平衡(虛線)所隔開。
1. 穩態轉化的三維圖示
生態系統穩定狀態之間的轉化可用三維圖(圖1)形象地表示,這一穩定性景觀刻畫了在五個不同條件(如營養鹽水平)下的生態系統的狀態(如濁度)及其吸引域。穩定平衡對應于波谷,而摺疊的位於中段的不穩定平衡對應于波峰。如果吸引域的容積較小,即使一個中等程度的擾動都有可能將系統推進到另一個吸引域中去。
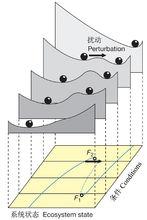 圖1 “杯中彈子”圖示五個不同狀態生態系統的穩定狀態
圖1 “杯中彈子”圖示五個不同狀態生態系統的穩定狀態2. 穩態轉化的化學計量機制
淺水湖泊中由於化學元素輸入失衡導致的災變性穩態轉換之所以難以預測,主要是因為缺乏對相關機制的清晰了解。多穩態理論認為,富營養化對淡水生態系統的結構、功能和穩定性有著明顯的負面影響,但是,人們仍然不清楚在一個突變點出現之前富營養化是如何通過化學計量效應使生態系統去穩定化的。生態化學計量學的關鍵參數——化學計量內穩性(stoichiometric homeostasis,簡稱 H)連結小尺度過程與大尺度格局之間的橋樑,可用於揭示湖泊生態系統的穩態轉換(清水⇄濁水)機制。
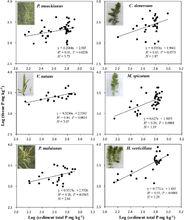 圖2沉積物P與沉水植物地上組織中P含量之間的關係
圖2沉積物P與沉水植物地上組織中P含量之間的關係圖2 表示沉積物P與優勢沉水植物種類地上組織中P含量之間的關係。化學計量內穩性係數(H)根據公式log(y) = log(c) + (1/H)log(x)計算所得,其中y是植物的P含量,x是沉積物的P含量,c是常數。左邊三種為高內穩性植物——分別是微齒眼子菜( Potamogeton maackianus)、苦草( Vallisneria natans)和馬來眼子菜( P. malaianus),右邊三種為低內穩性植物——分別是金魚藻( Ceratophyllum demersum)、穗狀狐尾藻( Myriophyllum spicatum)和輪葉黑藻( Hydrilla verticillata)。
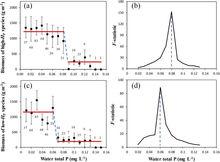 圖3高內穩性/低內穩性沉水植物生物量與水中總磷的關係
圖3高內穩性/低內穩性沉水植物生物量與水中總磷的關係基於對長江流域97個淺水湖泊的調查,通過測量常見沉水植物地上組織和沉積物中的氮(N)、磷(P)濃度發現,沉水植物存在顯著的磷化學計量內穩性P ( H)而非氮N ( H),閾值檢測(Threshold detection)表明,高內穩性種類(微齒眼子菜、苦草和馬來眼子菜)占優勢的沉水植物群落髮生穩態轉化的臨界磷濃度較高(0.08 mg/L),而低內穩性種類(金魚藻、穗狀狐尾藻和輪葉黑藻)占優勢的沉水植物群落出現穩態轉化的臨界磷濃度較低(0.06 mg/L),表明高內穩性沉水植物對富營養化的可塑性更強(圖2)。隨著富營養化的發展,低內穩性植物先行崩潰,這可作為湖泊生態系統從清水到濁水穩態轉化的早期預警信號。但低內穩性植物具有較快的恢復能力,因此可作為生態修復的先鋒物種(圖3)。
圖3 表示高內穩性(high H)/低內穩性(low H)沉水植物生物量與水體中總磷之間的關係,並分別對兩種類型沉水植物進行了變點分析。黑點與縱線表示湖水每0.01 mg/L TP間隔中的平均水生植物生物量和標準差。紅線表示突變閾值前後水生植物的平均生物量。虛線表示閾值的位置,在標準差上/下的數字表示每個間隔的樣本數。
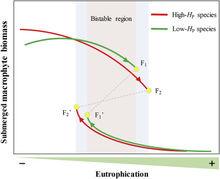 圖4解釋淺水湖泊穩態轉化機制與過程的模式圖
圖4解釋淺水湖泊穩態轉化機制與過程的模式圖圖4 解釋淺水湖泊穩態轉化機制與過程的模式圖。與由低內穩性物種占優勢的生態系統相比,高內穩性沉水植物占優勢的生態系統具有較高的生產力、穩定性和可塑性(由高營養鹽閾值所表征)。高內穩性植物占優勢的生態系統具有較寬的雙穩態區,在前行衰退軌跡的末端具有較低的生物量,但由於保守的營養鹽利用策略和較低的生長速率,在沿著恢復軌跡趨於平衡時具有較高的豐度。
沉水植物P的化學計量調節能力不僅是預測生態系統結構和穩定性的強有力的指標,也與在面臨外界干擾下物種演替與生態系統可塑性變化相關聯。高內穩性植物占優勢的生態系統傾向於具有更穩定的狀態,而低內穩性植物占優勢的生態系統更為機會主義,對外界的擾動更為脆弱。簡言之,富營養化通過首先抑制高內穩性的沉水植物群落來使湖泊生態系統去穩定化,但低內穩性的沉水植物群落由於較低的突變閾值而先行崩潰,這恰好可用作湖泊生態系統從清水到濁水轉化的早期預警信號。但同時由於低內穩性植物具有較快的恢復能力,因此可作為生態修復的先鋒物種。化學計量內穩性在決定沉水植物占優勢的湖泊生態系統的結構、穩定性和可塑性中扮演的角色具有重要意義。在全球富營養化日趨嚴重的形勢下,穩態轉化的化學計量機制對湖泊生態系統的管理與恢復具有重要的理論與實踐意義。
3. 穩態轉化成功案例
 圖5. 生態修復前的錦城湖——藍藻飄蕩,水色混濁
圖5. 生態修復前的錦城湖——藍藻飄蕩,水色混濁位於成都市區的錦城湖1號湖,湖區水域面積約11.3 萬平方米,原是劣五類水質,水生植物消失殆盡,每逢夏季,湖面漂浮著一層像綠油漆一樣的藍藻水華,呈現典型的濁水狀態(圖5)。
在成都市政府的資助下,四川中科水務以穩態轉化理論為基礎,通過標本兼治的方式,對嚴重受損的湖泊生態系統進行了修復與最佳化,近一年的時間,就建立了一個物種豐富、結構完整、動態平衡的清水生態系統(圖6)——恢復了水生植物的多優群落(苦草、微齒眼子菜、馬來眼子菜、狐尾藻、黑藻等),沉水植物覆蓋度高達95%,形成了一片壯麗的水下森林景觀,清澈見底(透明度達4.5m),水質基本處於I類到II類的水平,達到集中式生活飲用水地表水源地一級保護區的水質標準。
 圖6.生態修復後的錦城湖——水下森林,清水穩態
圖6.生態修復後的錦城湖——水下森林,清水穩態