
pSV2及其派生的質粒載體
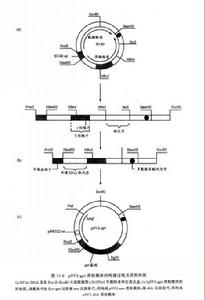 圖1
圖1大腸桿菌的Eco—gpt基因已經被克隆出來,並插入到各種不同的載體上,例如SV40晚期取代載體pSV2,構成新的重組質粒載體pSV2一gpt,使其置於SV40啟動子的控制之下進行表達,因此有些作者也叫這種質粒載體為表達載體(圖1)。猿猴細胞在感染上了這種質粒載體之後,便獲得了表達Eco—gpt基因的能力,而且依據其對次黃嘌呤抑制作用的不敏感性和電泳遷移率的差別,就可以把這種外源的Eco—gpt表達產物,同內源的哺乳動物之HGPRT區別開來。
pSV2是在DNA腫瘤病毒SV40的基礎上發展出來的一種新型的質粒載體家族。它是由從SV40早期轉錄單位派生出來的一種表達盒(expression cassette),插入在pBR322質粒DNA上重組形成的。這個表達盒共有三個組件。第一個組件是Pvu Ⅱ—Hind Ⅲ區段,它含有SV40早期轉錄啟動子及轉錄起始位點,但沒有轉譯起始密碼子ATG。這個組件的功能是啟動插入在它的下游的外源基因進行轉錄;第二個組件是含有66bp長的小t蛋白質間隔子的DNA區段,其作用在於為異源的轉錄本提供一種剪輯給體及受體的位點;第三個組件是含有轉錄終止區及多聚腺苷酸化位點的DNA區段,起到終止轉錄反應和poly(A)加尾的功效。
pSV2質粒載體中另一種重要組成部分是pBR322質粒的DNA序列,包括一個原核的複製起點,和一個氨苄青黴素抗性(Amp)選擇標記。
外源的選擇標記基因Eco—gpt,以及neo和dhfr,以保持正確讀碼結構的Hind Ⅲ—Bgl Ⅱ片段形式,定向插入到pSV2質粒載體的Hind Ⅲ和MboI克隆位點上(Bgl Ⅱ和MboI為同尾酶),由此便產生出pSV2一gpt、pSV2一neo和pSV2一dhfr三種不同的質粒載體,其中大T間隔子序列被破壞掉了(圖1)。這些質粒載體在哺乳動物基因工程中有著十分廣泛的用途,為克隆的異源基因提供了便利的檢測、選擇及表達的手段。
pRSV及其派生的質粒載體
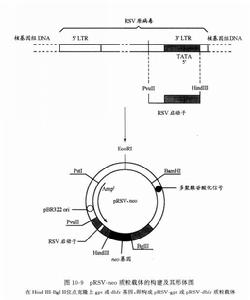 圖2
圖2在發展出了上述這些以SV40啟動子為基礎的表達載體之後,科學工作者又發現在某些哺乳動物的細胞中,勞斯肉瘤病毒(Rous sarcoma virus,RSV)啟動子比SV40的具有更強的表達能力。RSV是一種單鏈RNA反轉錄病毒,它是經由雙鏈DNA形式整合到寄主染色體上。RSV的啟動子便是位於此種原病毒的長末端重複序列(LTR)中。現已分離到含有RSV啟動子的長度為524bp的DNA片段。用此片段取代pSV2質粒載體中的Pvu Ⅱ—Hind Ⅲ(SV40啟動子)區段,便構成了哺乳動物細胞的另一類表達質粒,即pRSV質粒載體系列,包括pRSV—gpt及pRSV—neo等(圖2)。可能是由於在強啟動子作用之下,這類質粒載體上的選擇標記基因獲得了更加有效的表達。因此套用此類質粒載體,顯然可以得到相對較高的DNA轉染效率。
質粒載體
質粒DNA是微生物細胞中分子量比染色體DNA小得多的共價、閉合、環狀雙鏈DNA分子(個別除外,如酵母自殺質粒是RNA),是一種存在於染色體外的能自主複製的遺傳因子,質粒通常帶有與細胞的主要代謝活動無關的一些基因,例如抗生素抗性基因、產生細菌素的基因、糖類分解代謝的基因和誘發腫瘤的基因等等。由於質粒的存在,宿主細胞往往被賦予新的表型,當把一個含抗藥性基因的質粒轉入細胞之後.原來無抗藥能力的細菌則表現出抗藥新表型。
質粒的生物學性質:
(1)質粒的複製具備複製起點(原點)是DNA分子在宿主細胞中進行複製的一個必要條件。根據複製控制類型的不同,質粒分為嚴緊型質粒與鬆弛型質粒,前者受宿主細胞複製作用的嚴格控制,因此,每個細胞中只含有一至幾個拷貝;而後者則受宿主細胞的控制不嚴,它們在每個細胞中的數目可達10—500個拷貝,當用氯黴素抑制細胞蛋白質合成時,質粒拷貝數可擴增至數千個。現在使用的質粒載體絕大多數都是鬆弛型質粒。
(2)質粒不相容性質粒不相容性(incompatibility)指在沒有選擇壓力的條件下,兩種親源關係密切的質粒不能共存於同一宿主細胞中的現象。原因是它們的複製子相同,所用的複製系統也相同,故在複製和分配到子細胞的過程中互相競爭。細菌生長几個世代後,量少的質粒就完全消失。至今已發現30個以上的不相容組,只有屬於不同不相容組中的質粒才能共同存在於同一個宿主細胞中。
(3)選擇標記 當質粒DNA轉化宿主細胞時,只有極少部分宿主細胞接受了DNA,所以需要利用質粒編碼的選擇標己將轉化成功的宿主細胞(陽性克隆)從大量的宿主細胞巾篩選出來。
質粒最常用的選擇標記是抗生素抗性基因,這些抗性主要包括氨苄西林(Amp或Ap)、四環素(Tet)、氯黴素(Cm)、卡那黴素(Kan或Km)和新黴素(Neo)等,例如,對氨苄西林有抗性的質粒,一般寫作Amp;而對該藥敏感的質粒則寫作Amp。
含有氨苄西林抗性質粒的細菌之所以對氨苄西林產生抗性是由於氨苄西林抗性基因的編碼產物是β一內醯胺酶,該酶存在於細菌的細胞膜與細胞壁之間,可將細菌周圍的氨苄西林降解,從而使細菌生存下來、值得注意的是:抗生素抗性基因必須完整,才能表達出有活性的酶去破壞抗生素而使宿主菌表現出抗藥性,如果把抗性基因切斷、分開或改變,如在其中插入一段外源基因,則抗性基因便無法表達出有功能的酶而使抗生素失活,這種現象就叫做插入失活。利用插入失活可使細菌由耐藥變為敏感的特點,可以對質粒是否插入了外源基因進行快速篩選。
