![洄游[一種魚類運動]](/img/0/71b/nBnauM3XzczMxAzM5ADN1QDM1UTM1QDN5MjM5ADMwAjMwUzLwQzL0EzLt92YucmbvRWdo5Cd0FmLxE2LvoDc0RHa.jpg "洄游[一種魚類運動]")
洄游類型
魚類洄游有不同的劃分方式。按洄游的動力,可分為被動洄游和主動洄游;按洄游的方向,可分為向陸洄游和離陸洄游、向河(海)洄游和溯河洄游等;根據生命活動過程中的作用可劃分為生殖洄游、索餌洄游和越冬洄游。洄游是一些魚類的基本屬性,如果不能完成洄游或洄游受到阻礙,則這些魚類的生命周期將遭到破壞,並影響群體的增殖,甚至危及物種的生存。
索餌洄游
與生殖洄游和越冬洄游不同,有的魚類在洄游的過程中就往往達到了洄游的目的。索餌洄游的特點是洄游路線、方向和時間隨著餌料生物群的分布和密度,索餌魚群數量多寡和狀態而變動。
索餌魚群較不穩定,也較分散。索餌期的長短主要取決於餌料生物的數量和質量,魚的豐滿度、含脂量以及水溫狀況等。
生殖洄游
又稱產卵洄游。魚類的產卵習性是多種多樣的,有的魚為了尋求適宜的產卵條件,保證魚卵和幼魚能在良好的環境中發育,常常要進行由越冬場或肥育場向產卵場的集群移動。洄游距離有長有短,因種類而異。生活在淡水中的鰻鱺,要洄游數千公里到海洋深處產卵,而生活在大洋中的大馬哈魚要反向遊動數千公里進入江河上游產卵。而有些魚類僅作短距離移動,如大黃魚由福建北部的洞頭洋洄游至江蘇海域的呂泗洋;中國淡水中的一些鯉科魚類,如青魚、草魚、鰱、鱅等魚類由靜水湖泊洄游至江河中特定河段的產卵場所產卵,距離長則幾百公里,短則幾十公里。
生殖洄游的移動方向依種類而異,有由深海、外海游向淺海或近岸;有由海洋向江河或江河下游游向上游的溯河洄游;有由江河向海洋的降河洄游;湖泊中的魚類由開闊的湖中心區游向沿岸產卵,也可以視作是一種產卵洄游,但洄游的距離很短。生殖洄游的特點是洄游路線、方向和時間較固定,距離較遠,游速快,分群洄游現象明顯,魚群密度大,魚群集群性狀固定;洄游期間魚體性腺發育、生理狀態變化大。有些魚類體型、體色出現變異。有些魚類在生殖洄游過程停止攝食。
越冬後的幼魚和成魚,經過生殖洄游或生殖活動的成魚,需要強烈的攝食以補充營養,集群游向餌料豐富的水域索餌,形成了索餌洄游,也稱攝食洄游。有些魚類在越冬後先行強烈攝食,形成由越冬場向肥育場地的洄游,例如在河流深槽中越冬的鯉魚,在早春氣候轉暖時,就要洄游到食料豐盛的湖泊中去肥育。攝食洄游也有伴隨產卵洄游同時進行的,如鮪魚在由西沙群島進行長距離生殖洄游過程,沿途強烈攝食,實際上鮪魚的生殖洄游也是一種索餌洄游。
有些魚經過生殖後,在秋季進行一次索餌洄游,為準備越冬和來年生殖積儲營養。幼魚的攝食洄游也是十分明顯。如草魚、青魚、鰱、鱅等魚類在江河繁殖後,已經發育到能自由游泳和自由攝食的幼魚,即沿岸逆水上溯,進入通江的湖泊或小支流中進行肥育,因為這些附屬水體中的食物要比江河幹流中豐盛得多。
洄游魚類
由於魚類棲息環境的不同,洄游性質的不同。人們常常把洄游魚類分成幾大類。常用的分法為:海洋洄游魚類、淡水洄游魚類、河口半鹹水洄游魚類、過河口性洄游魚類等。根據魚類洄游的性質,把進行長距離移動和海河之間運動的魚類稱作洄游魚類,把不同淡水水體之間的移動或江河上下游之間的移動稱作半洄游魚類。
海洋魚類的洄游
完全在海洋中生活和洄游,同種魚往往分成若干種群,每一種群有各自的洄游路線,彼此不相混合,各海區的魚群有不同的變異特徵,每個海區都分布有它自己的洄游群體。如中國東海、黃海的小黃魚可分為4個種群,分別有其自己的越冬、產卵與索餌的洄游路線。
溯河性魚類的洄游
溯河性魚類生活在海洋,但溯至江河的中上游繁殖。這類魚對棲息地的生態條件,特別是水中的鹽度有嚴格的適應性。如北太平洋的大麻哈魚溯河後即不攝食,每天頂著時速幾十千米的水流上溯數十千米,在洄游過程中體力消耗很大,到達產卵場時,生殖後親體即相繼死亡。幼魚在當年或第二年入海。但某些生活在河口附近的淺海魚類,生殖時只洄游到河口,如長江口的鳳鱭等,溯河洄游的距離較短。
降海性魚類的洄游
降海性魚類絕大部分時間生活在淡水裡而洄游至海中繁殖。鰻鱺是這類洄游的典型例子。歐洲鰻鱺和美洲鰻鱺降海後不攝食,分別洄游到數千千米海域後產卵,生殖後親魚全部死亡。其幼魚回到各自大陸淡水水域的時間不同,歐洲鰻鱺需3年,美洲鰻鱺只需1年。中國的鰻鱺、松江鱸等的洄游也屬於這一類型。海、淡水鹽度不同,滲透壓有差異,因此作溯河或降海洄游的魚類,過河口時往往需要在鹹淡水區停留一段時間,以適應這種生理機能的轉變。
半洄游魚類
半洄游魚類是指淡水魚類在淡水水域中從一種類型的水體到另一種類型的水體,以及棲居在淺海區或鹹淡水區的魚類進入江河的洄游。歸納起來,主要可以分為兩種類型,即江湖半洄游和河口乾流半洄游。
中國著名的四大家魚草魚、青魚、鰱、鱅等都是半洄游魚類。這些魚類平時在江河幹流的附屬湖泊中攝食肥育,繁殖季節結群逆水洄游到幹流的各產卵場生殖。產後的親魚又陸續洄游到食料豐盛的湖泊中索餌。棲息在江河下流的親魚魚群也有洄游到中游或上游產卵場產卵。它們在洄游的過程中,性腺逐漸達到成熟。江河半洄游魚類,秋季在湖泊中進行覓食肥育後,秋末冬初進行越冬洄游,即從較淺的湖泊中游到江河幹流的河床深處越冬。但這些魚類的越冬洄游有時不很明顯,當湖泊中也存在適合越冬的深水區時,如深窪或潭坑等,也可以在這些場所越冬。翌年開春,水溫上升,這些在江河幹流或湖泊深水區的坑、潭中越冬的魚類又重新洄游進入湖泊索餌。
江湖半洄游魚類的幼魚,半洄游習性也非常顯著和有規律性。在出生的當年夏季,當它們已具備自由游泳的能力後,即沿岸溯游作攝食洄游。在支流的河口或湖泊的通江泄水口處,可以見到幼魚魚群奮力克服較大的流速,進入支流或湖泊中覓食。幼魚這種克服流速的能力,除了魚類自身具有逆流運動的特性和幼魚的強烈索餌要求之外,水流中的食物逐漸豐富可能是十分重要的外界條件。幼魚雖與成魚一樣,具有非常顯著和有規律性的索餌洄游習性,但幼魚沒有明顯的越冬洄游和生殖洄游。幼魚在秋季完成索餌洄游後,在秋末冬初由湖泊淺水區移動到深水區越冬,一般不到江河幹流越冬。翌年開春水溫回升後,由湖泊深水區游向沿岸淺水區餌料生物豐富的區域索餌、生長、肥育。
半洄游魚類中另一類型為河口乾流半洄游魚類。這些魚類平時生活在河口或半鹹淡水區,生殖季節進入江河,上溯至中、下游的適合場所生殖。如鱭屬魚類是典型的河口乾流半洄游魚類。長頜鱭(又稱刀鱭)平時分布於長江及近海半鹹淡水區,生殖季節從河口進入淡水,沿幹流上溯至長江中游產場作生殖洄游,最遠可達洞庭湖,有的在江河幹流產卵,也有進入支流及通江湖泊產卵。產卵後親魚分散在淡水中攝食,並陸續緩慢地順流返回河口及近海,繼續肥育。長頜鱭的幼魚也順水洄游至河口區肥育。冬季,長頜鱭不作遠距離洄游,而聚集在近海深處越冬。
洄游魚類與半洄游魚類的洄游性質沒有差別。它們不同的地方是洄游魚類對產卵場有較嚴格的要求,洄游的距離較遠,繁殖時期較為集中。但這種區別不是絕對的,有些魚類的洄游習性界於洄游和半洄游之間。有些定居型的魚類,由於對環境的適應性較強,大多不進行明顯的洄游或半洄游運動,但在環境許可的條件下,如鯉魚也可以進行湖河越冬洄游,春季又進行河湖攝食洄游。
生態原因
產生魚類洄游的原因,是由於魚類本身的生理要求,包括對餌料豐富水域、適宜的產卵地或越冬場所的追求。影響魚類洄游的環境因子有水流、底形、溫度、鹽度、水質、光線等,其中水流是對洄游的定向起決定性作用的因子。在具有一定流速的條件下,魚類通常都逆流而游。
進行生殖洄游的魚類,往往隨即進行索餌洄游,有的在冬季降臨時進行越冬洄游。但是,並不是一切洄游魚類和半洄游魚類都要進行所有的三個洄游環節,有些冷水性魚類,如白鮭只進行生殖洄游和索餌洄游,沒有越冬洄游。白鮭多在晚秋和初冬產卵,冬季也不停食。生活在淺水湖泊的一些魚類,攝食和產卵都在湖區內進行,有時只進行游向河床深處的越冬洄游。幼魚則只有攝食洄游,冬季依舊攝食,不存在越冬洄游。
啟動誘因
魚類洄游的起始,既取決於魚自身的狀態,也取決於周圍環境條件的影響。例如魚性腺成熟所分泌的性激素刺激神經系統興奮而產生生殖要求。環境條件的變化則是開始生殖洄游的天然刺激信號。溫帶地區達到一定豐滿度的魚,溫度下降的天然刺激就成為開始越冬洄游的信號。
多數魚類如果性腺發育不良,即使已經達到生殖年齡,外界環境刺激強烈,仍不會產生生殖洄游的要求。沒有達到一定豐滿度或含脂量的魚,儘管外界水溫下降劇烈,仍繼續索餌或邊洄游邊索餌。如歐洲鯷魚,當含脂量為14%以下時,即使水溫劇降,越冬季節已經到來也不開始洄游,而是繼續攝食。當含脂量為14-17%時,才開始洄游,但行動緩慢。含脂量達到22%時,到了越冬季節,則不論水溫下降幅度的大小,就立即開始越冬洄游。環境條件中以水溫、水流、水化學等的影響最為顯著。水溫回升快,開始生殖洄游就早。水溫的變化會影響餌料生物的發生和分布,從而影響魚類的索餌洄游。秋末水溫下降的遲早與快慢直接影響越冬洄游的開始時間和洄游速度。水流是造成被動洄游和主動洄游的主導因素。水流把魚卵和仔、稚魚攜帶遠離出生地,形成成魚回歸性洄游。水的化學成分,特別是鹽度的變化,引起魚體內滲透壓的改變,導致血液成分與性質的變化,進而誘發魚的生理變化,促使降海或溯河洄游的發生。
形成機制
洄游是魚類在漫長的進化歲月里自然選擇的結果,通過遺傳而鞏固下來。魚類洄游具有定期性、定向性、集群性和周期性等特點。幾乎所有的洄游都是集群洄游,但不同種類不同性質的洄游,洄游的集群大小各不相同,這與保障最有利的洄游條件有關。洄游距離的遠近與洄游魚類的體型大小及其自身狀態有關。體型大,含脂量高,洄游距離較遠,如鱘、大麻哈魚、鰻鱺等的行程均達數千里。洄游的定向性除與遺傳性有關外,高靈敏度和選擇性的嗅覺,在引導鮭、鰻鱺等魚類數年之後歷程數千公里回歸原出生地起了很大作用。鮪魚的顱骨內極其細小的磁粒,使其在大洋中洄游不會迷失方向。側線靈敏的感流能力也起著引導洄遊方向的作用。
脅迫因子
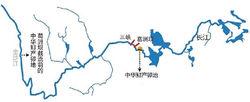 圖1 長江中華鱘產卵示意圖
圖1 長江中華鱘產卵示意圖洄游是在漫長的生命演化與適應過程中形成的秉性,難以在人類活動的時間量程內進行根本性改變,因此,人類干擾(如修築大壩)容易引發一些物種的絕滅。譬如,20世紀70年代,長江中的中華鱘繁殖群體尚有1萬餘尾,1981年葛洲壩建成後,阻止了中華鱘親魚的生殖洄游,使親魚無法上溯到金沙江產卵(圖1),中華鱘繁殖群體開始急速衰退:1983-1984下降到約2176尾,2005-2007年期間下降到了203-257尾,到了2010年只剩數十尾。中華鱘已被列入《世界自然保護聯盟》(IUCN)2012年瀕危物種紅色名錄ver 3.1——極危(CR)。
