概述
染色體遺傳學說,是美國生物學家摩爾根對孟德爾學說的進一步發展。他用果蠅做了大量試驗,提出了遺傳因子位於染色體上,並稱這些因子為基因。染色體存在於細胞核中,在細胞期(細胞不分裂的時期),它以染色質的狀態存在;在細胞分裂時,成為短粗的桿狀結構,稱為染色體(因為它染色較深)。染色體實際上是染色質濃集而成的,內部呈緊密的、高度螺鏇曲卷的絲狀結構。在細胞核中含有進一步發育所必需的所有信息,它決定這個細胞將發育成為一個人、一匹馬或一羽鴿子,並且決定鴿子是大型或小型的,羽色是灰的或是紅的,飛翔能力是優秀的或是低劣的等。這些發育或發展的資料及指令,均存在於染色體的絲狀結構上。在染色體上,依照順序包含一系列鹼基,稱為基因。基因在染色體上的分布,就好像項鍊上成串的珠子,這些成串的基因正代表著所有的遺傳性狀,因而被稱為“遺傳的基本單位”。
簡介
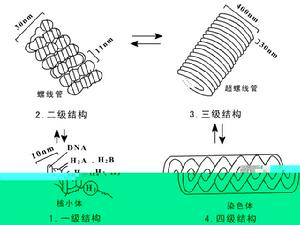 染色質
染色質在細胞分裂之前,所有細胞內的基因、染色體及細胞核等行將分離的部分,都會由一個分裂成為平均的兩個部分。成對的染色體,在科學上稱為同源染色體。同源染色體中的兩條染色體形狀及大小都相同,但兩者之間並非是完全一致的。成對的同源染色體中的1條有時可能比另1條多出1個或1個以上不同的基因。這種1個基因的2價形式稱為對偶基因。
在一個個體細胞中的一個同源染色體對中,其中一條是來自雄性親代,而另一條則來自雌性親代。因此,每一個性狀均由兩個基因所控制,其中一個來自雄性親代,另一個來自雌性親代。如果兩個基因完全相同,那么這對基因為同合子,如果同屬於一個對偶基因而不相同,便稱為異合子,或非純合子。
一個異合子基因對可以不同的方式去影響個體的性狀表現。如果其中的某一基因的表現強過另一個基因,那就整對基因而言,將會表現這個強勢基因效果,形成顯性基因及隱性基因。如果用純種的黑色公牛和純種的紅色母牛雜交,其子代的毛色都是黑色者,那么前者就是顯性基因,而後者則為隱性基因。然而,在第一子代(Fl)的染色體中,隱性基因雖受到顯性基因的壓制而沒能有所表現,但是它仍存在於其染色體中。而且在第一子代彼此間雜交時,則將在凡子代中表現出來,而使几子代個體中,出現紅色的個體,這就證明隱性基因並沒有消失,而只是被壓制。在其他形式的雜交配種時,也可能發生子代毛色的兩個親代都不同,而是介於兩個親代的毛色之間,成為一種兩個親代混合型。
建立過程
德爾豌豆雜交試驗論文被重新發現3年後的1903年,薩頓(Sutton)和鮑維里(Boveri)根據各自的研究,認為孟德爾“遺傳因子”與配子形成和受精過程中的染色體行為具有平行性,同時提出了遺傳的染色體學說,認為孟德爾的遺傳因子位於染色體上,這個從細胞學研究得出的結論,圓滿地解釋了孟德爾遺傳現象。
9世紀,特別是下半葉,細胞學有了空前的發展:魏斯曼(A.Weismann)首先提出要在細胞水平研究遺傳的觀點,斷然否定獲得性遺傳,提出種質連續理論;赫特維希(O.Hertwig)研究海膽受精作用,確認動物細胞在形成配子過程中要發生減數分裂;魯(W.Roux)1883年提出遺傳物質均等分配的觀點;薩頓和蒙哥馬利(Montgomery)證明了染色體的連續性,薩頓指出:“父本與母本的染色體以配對的方式聯繫,減數分裂時彼此分開,這就是孟德爾遺傳的物質基礎”;鮑維里研究馬蛔蟲受精作用,證實了染色體的個體性,他認為:從受精卵衍生出來的一切細胞,其中的染色體一半來自父方、一半來自母方。這正如威爾遜(E.B.Willson)所預言的:“細胞通過分裂在發生上連續,遺傳現象就是這種連續性的結果。”
這些細胞學成果的染色體假說在1903~1910年的這段時間內未得到承認和支持,有不少遺傳學家反對,貝特森(Bateson)就屬其中之一。連鎖現象最初是貝特森於1906年在香豌豆雜交中發現的,儘管此時染色體學說已經提出3年,但他在連鎖現象面前,除了提出相斥和相引這兩個名詞和“珍惜你的例外吧!”這句名言之外,別無建樹。
後來摩爾根對染色體學說作出了決定性貢獻。
染色體理論主要來自細胞學的觀察事實,摩爾根認為這是“思辨”臆測,要求提供更直接的證據,宣稱絕不接受這種“沒有實驗基礎”的理論。
在摩爾根看來,承認那種性狀由“顆粒”控制,而顆粒位於單個不同的染色體上,這有餑於他的漸成論思維背景,這意味著將在一定程度上否定自己原有的概念體系。現在看來可以這樣認為:1910年之前,作為胚胎學家的漸成論者,對細胞學家所固守的預成論和顆粒論是持否定態度的,儘管預成論與漸成論的爭辯表面上看,100年前就已偃旗息鼓,且以後者勝利而告終,但胚胎學家對預成論思維的任何痕跡都過度敏感,因為對於預成論者,孟德爾學說在一定程度上就是顆粒學說,而摩爾根則認為預成論全然否定了細胞質和環境的作用。
當然,原初的顆粒遺傳理論確有簡單化的缺點。1909年詹森(W.Johannsen)才提出基因型和表現型的概念,在這之前,顆粒論者就不得不從遺傳因子與生物性狀之間一一對應聯繫的預成論思維去探討遺傳。也就是說,雖然小人學說早已無人問津,但基因型就是縮小了的表現型的跡痕卻未能動搖。
為什麼摩爾根很快又成為染色體學說的主要倡導者和一系列有力證據的提供者呢?1904年美國細胞學家威爾遜邀請他到哥倫比亞大學,作為好友、同事,兩人的辦公室僅一牆之隔,且有著一些既研究果蠅遺傳又研究昆蟲染色體的共同學生作媒介,此間威爾遜的工作深刻地影響著摩爾根。
1908年摩爾根引進黑腹果蠅建立果蠅室,作為遺傳學實驗材料。1910年5月發現一種白眼雄蠅,他立即用白眼雄蠅與正常紅眼雌蠅交配,結果F1全是紅眼,說明白眼屬隱性突變性狀;在F1兄妹交產生的F2中,出現了白眼只限於雄蠅的現象,這預示眼色與性別的遺傳相關;他又將這隻白眼雄蠅與F1紅眼雌蠅回交,結果表明雌蠅也可出現白眼;再將這隻白眼雌蠅與紅眼雄蠅雜交,出現了雄蠅全是白眼而雌蠅全是紅眼現象,他將這種女象父、子象母的現象稱為交叉遺傳(criss-crossinheritance)。
性狀遺傳隨性別不同而有所差異,這是孟德爾雌雄同株的豌豆不可能表現的新現象,但這使摩爾根聯想到世紀之交的性染色體研究。
1891年後亨金(H.Henking)發現並命名半翅目昆蟲精巢細胞中含異染色質的X染色體;1902年麥克隆(C.E.Mcclung)將該染色體與昆蟲性別相聯繫;威爾遜(1905年)研究半翅目和直翅目昆蟲,認為性別由性染色體決定,特別是史蒂文斯(N.M.Stevens)發現果蠅性別決定機制。
運用這些細胞學成果,可斷言上述基因位於x染色體上,但摩爾根仍然躊躇不定,其主要原因是:漸成論觀點在複雜的實驗結果面前,仍影響著摩爾根對並非基於“小人與活力”之上的預成論觀點的思維。他很難相信染色體是一種能夠從根本上控制一切的“顆粒”。
摩爾根果蠅雜交結果與英國研究的蛾類和鳥類性別決定機制相牴觸;後來證實這種平行的相反恰是基因位於染色體上的又一證據。
果蠅只有8條染色體,而個體性狀紛呈繁雜,他擔心這樣一來,許多性狀勢必包含在同一染色體中而隨之一道“孟德爾化”。物種間染色體數目差異甚大,這使他認為染色體未必是能控制一切性狀的基質。
縝密的科學思維,可貴的懷疑主義!當進一步的研究使他對這些問題有所醒悟時,著名的論文《果蠅的限性遺傳》誕生了,在遺傳學這是第一次將一個具體的基因與一個特定的染色體聯繫起來。
然而最終直接證明染色體理論的還是摩爾根的合作者布里吉斯的經典研究。他在重複白眼雌蠅與紅眼雄蠅的交配時,發現了約1/2000的初級例外子代,即女偏母、子偏父的“性狀直傳”;再將初級例外中白眼雌蠅與正常紅眼雄蠅交配,發現了更高頻率(4%)的次級例外。布里吉斯假設初級例外的白眼雌蠅具有XXY的異常性染色體(即母本減數Ⅰ過程中X染色體不分離,共同進入受精卵所致),它形成的4種配子與正常父本的2種配子結合,即形成8種眼色、性別、育性、生活力不同的後代,其中4%為直傳性狀的次級例外,布里吉斯還逐一鏡檢了這些果蠅的性染色體,結果完全證實了他的假說,同時還提出了性別平衡理論。這項研究直觀地、令人信服地證實了染色體就是基因的載體。
遺傳的染色體學說的誕生是細胞學和遺傳學的結晶,宣告了細胞遺傳學的日臻成熟,正如米勒所言:“它為整個現代遺傳學奠定了基礎養料”。摩爾根也因此在諾貝爾誕生100周年的1933年而榮膺桂冠。諾貝爾委員會認為:“沒有摩爾根的研究,就沒有人類遺傳學,也就沒有人類優生學”。遺傳學的飛速發展業已表明,當時的這種評價遠遠不夠。正如科學史學家邁爾(E.Mayr)1984年所言:“接受遺傳的染色體理論絕不是染色體研究的終止,而是進入染色體研究新時代的標誌”。
