概述
 你
你數量性狀(quantitative characters)是指在一個群體內的各個體間表現為連續變異的性狀,如動植物的高度或長度等。數量性狀較易受環境的影響,在一個群體內各個個體的差異一般呈連續的常態分配,難以在個體間明確地分組。生物界的另一類性狀如紅與白、有與無等稱質量性狀。質量性狀比較穩定,不易受環境條件的影響,它們在群體內的分布是不連續的,雜交後代的個體可以明確地分組,因而可以計算雜交子代各組個體數目的比率,分析基因分離、基因重組以及基因連鎖等遺傳行為。
數量性狀在生物全部性狀中占有很大的比重,一些極為重要的經濟性狀(如作物產量、生育期、籽粒重、乳牛泌乳量、羊毛長度等)都是數量性狀。研究數量性狀遺傳規律的學科稱為數量遺傳學。
特徵
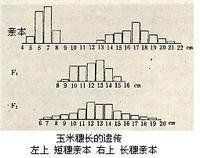
①個體間的差異是連續的,例如用穗長有差別的兩個玉米品種進行雜交,則子一代(F1)植株的穗長介於兩親本之間,子二代(F2)植株的變異幅度擴大,子代各個植株的穗長呈連續的變異,因而無法求出穗長的分離比率而只能用一定尺度測量性狀的表型值,再用統計學方法加以分析(見圖);
②容易受環境的影響,甚至純合的親本或基因型一致的子一代的表型也呈現連續變異。所以子二代的變異一方面來自基因重組,另一方面則來自環境的影響。
多基因學說
多基因學說於1909年由瑞典學者H·尼爾松·埃勒提出,他認為根據質量性狀研究的結果得來的孟德爾定律同樣可以用來解釋數量性狀的遺傳。多基因學說的要點是:①同一數量性狀由若干對基因所控制;②各個基因對於性狀的效應都很微小,而且大致相等;1941年英國數量遺傳學家K·馬瑟把這類控制數量性狀的基因稱為微效基因,相應地把效應顯著而數量較少的控制質量性狀的基因稱為主效基因;③控制同一數量性狀的微效基因的作用一般是累加性的;④控制數量性狀的等位基因間一般沒有明顯的顯隱性關係。
按照多基因學說,如果控制某一數量性狀的基因對數是N,則雜交子二代中該性狀表型的分布可以用二項式分布(1/2+1/2)2N展開的各項係數表示。例如小麥的種皮顏色由三對基因所控制,每一對基因的表型效應大致上相等而且是累加的,因此雜交子二代中出現的種皮顏色有七種,相當於二項式(1/2+1/2)2×3展開的各項。如果基因的對數更多,那么各組間的表型的差別將更小,分布也將更接近於常態分配。
數量性狀與質量性狀的關係
數量性狀與質量性狀的區分並不是絕對的。由於劃分的標準不同,往往也可以把數量性狀看作質量性狀。例如人的高脂蛋白血症是由多種基因決定的數量性狀,該症的某些生理、生化指標在人群中表現為連續的變異。但是從臨床的角度考慮則可以把人群劃分為患者和正常兩類,因而可以把它看作質量性狀。這種根據某一數量變化範圍來區分類別的數量性狀稱為閾值性狀。
決定數量性狀的基因也不一定都是為數眾多的微效基因。例如果蠅的突變型巨體(giant,gt)的個體明顯地比野生型大;小鼠的突變型侏儒(dwarf,df)的體型明顯地比野生型小;玉米的矮株基因(brachytic,br)同樣由於單個基因發生突變而使節間縮短,植株矮小。孟德爾的實驗中所採用的豌豆的高桿品系和矮桿品系同樣是一對主效基因的差別。此外,在特定條件下多對微效基因中的某一對的分離也可以使雜交子代中出現可以明顯地區分的表型。例如在水稻株高方面,當兩個品系的其他有關植株高矮的微效基因都相同而只有某一對基因不同的情況下,雜交子代中的植株高度便可以明顯地劃分為不重疊的高矮兩組。
數量性狀遺傳的多基因假說
在孟德爾遺傳規律被重新發現後的二十世紀初,形成了以BatesonW.和DevriesH.為首的Mendel學派以及以PearsonK.和WeldonW.F.R.為首的Galton學派,也稱為生物統計學派(Biometricians)。在遺傳和進化問題上,Mendel學派認為不連續性變異是重要因素,孟德爾原理可以普遍用於遺傳變異的研究,而連續性變異之所以不符合這些規律是因為它是不能遺傳的;而Galton學派則認為連續性變異是可遺傳的,是進化的重要因素,在研究上必須採用統計學的方法,而Mendel法則對於連續性變異不適用。這場爭論直到1909年才結束。該年詹森(JohannsenW.L.)發表了“純系學說(Purelinetheory)”,尼爾遜.埃爾(Nilsson-EhleH.)提出“多基因假說(polygenehypothesisormultiple-factorhypothesis)”;這兩個理論的建立,標誌著數量遺傳學的誕生。
