
基本概念
 蚜蟲的孤雌生殖(1)
蚜蟲的孤雌生殖(1) 很多種生物都有孤雌生殖現象的記錄,甚至在錘頭鯊這種較為原始的軟骨魚類身上竟也曾出現過孤雌生殖的現象。植物中黃瓜不經過傳粉受精就能結果。而在英國的切斯特郡立動物園在2006年5月期間,一隻單獨飼養的雌性科莫多龍flora居然發生了孤雌生殖的奇特現象——也算是稱得上是孤雌生殖現在在蜥蜴中的最高等的表現。
生殖類型
(一)偶發性孤雌生殖 (sporadic parthenogenesis):偶發性孤雌生殖是指某些昆蟲在正常情況下行兩性生殖,但雌成蟲偶爾產出的未受精卵也能發育成新個體的現象。常見的如家蠶、一些毒蛾和枯葉蛾等。
(二)經常性孤雌生殖 (constant parthenogenesis):經常性孤雌生殖也稱永久性孤雌生殖。這種生殖方式在某些昆蟲中經常出現,而被視為正常的生殖現象。可分為兩種情況 :
1、在膜翅目的蜜蜂和小蜂總科的-些種類中,雌成蟲產下的卵有受精卵和未受精卵兩種,前者發育成雌蟲,後者發育成雄蟲。
2、有的昆蟲在自然情況下,雄蟲極少,甚至尚未發現雄蟲,幾乎或完全行孤雌生殖,如一些竹節蟲、粉虱、蚧、薊馬等。
(三)周期性孤雌生殖(cyclical parthenogenesis):周期性孤雌生殖也稱循環性孤雌生殖。昆蟲通常在進行1次或多次孤雌生殖後,再進行1次兩性生殖。這種以兩性生殖與孤雌生殖交替的方式繁殖後代的現象,又稱為異態交替(heterogeny)或世代交替(alternation of generations)。如棉蚜從春季到秋末,進行孤雌生殖10一20餘代,到秋末冬初則出現雌、雄兩性個體,並交配產卵越冬。
生殖方式
(一)均等分裂型孤雌生殖:即卵原細胞正常進行減數分裂,產生3個極體和1個卵細胞,其中卵細胞獨立發育為後代個體的現象。(後代為單倍體)
(二)卵核與極體融合型孤雌生殖:即卵原細胞正常進行減數分裂,產生3個極體和1個卵細胞,其中卵細胞與任意極體隨機結合,形成“極體-卵細胞-受精卵”,並由此細胞發育成後代個體的現象(後代為2倍體)。
 蚜蟲的孤雌生殖(2)
蚜蟲的孤雌生殖(2) (三)分裂核融合型孤雌生殖:卵原細胞在進行減數第一次分裂時正常分裂,但不進行減數第二次分裂,最終形成了1個極體和一個“雙套卵”(diploid),由於它攜帶有母體的全套遺傳物質,自然可以獨立地發育為後代個體(後代為2倍體)。在一些偶發實例中,有的卵細胞形成後,因某種原因自然加倍,可以獨立發育為後代個體(後代也是2倍體),也算作這種情況。
(四)極體融合型孤雌生殖:即卵原細胞正常進行減數分裂,產生3個極體和1個卵細胞,但任意兩個極體間發生了融合,形成了“極體-極體”融合細胞,由於此細胞也攜帶有母體全套遺傳物質,也可以獨立發育為後代個體(後代是2倍體)。
以上四種情況在 蛾、蝶 中均有發現。另外,孤雌生殖有別於無性生殖,它是由生殖細胞而非體細胞完成的繁殖現象。二是產生的個體多數為單倍體,或者是進行重組之後的2倍體,而無性生殖產生的和母體遺傳物質完全相同的個體。所以通常把孤雌生殖歸類於有性生殖而非無性生殖。
相關區別
1.區別於無性生殖,是由生殖細胞而非體細胞完成的繁殖現象。
2.產生的個體多數為單倍體,或者是進行重組之後的2倍體,而非無性生殖產生的和母體遺傳物質完全相同的個體,所以通常把孤雌生殖歸類於有性生殖,而非無性生殖。
生殖起源
美國肯塔基大學生殖生物學博士後劉璟表示,首先應當問“為什麼有有性生殖”?她說,單細胞生物,比如細菌,是採用簡單的分裂方式繁殖的,它們絲毫不知道“兩性”的存在。低等生物比如水螅,可以通過“出芽”產生新的個體。這類無性生殖的方式多快好省,就好比人多長出一個手指來,然後手指自己掉下來,長成了一個小人兒,完全省去了“搞對象”的麻煩。那么,為什麼有性生殖會在演化中出現,並且成為演化的主流呢?
劉璟表示,現在大多數理論認為有性生殖能產生更多的基因組合,增加適應性演化的幾率,防止有害突變的積累。也就是說,有性生殖產生的多樣化的基因組合後代,其中一些後代能適應環境變化而生存下來;如果是無性生殖,後代基因總是一成不變,當環境發生改變時,所有後代可能都無法適應新環境。
最早的有性生殖的產生可能是因為有些細菌會融合在一起,然後再進行分裂。後來出現了兩個生物個體各出一個“配子”,然後配子結合形成新的生命。在二者的博弈中,生物形成了兩種完全不同的策略:一類生物提供具有極少營養物質的配子,也就是精子,可以支持“廣種薄收”;另一類生物提供具有大量營養物質的配子,也就是卵子,保證“成功率”。雄性和雌性的差別由此產生。
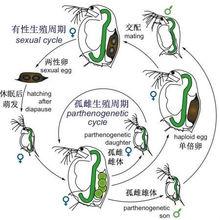 枝角類Daphnia的有性生殖與孤雌生殖的交替
枝角類Daphnia的有性生殖與孤雌生殖的交替 事實上,人們對有性生殖是如何起源知之甚少,從細胞遺傳學的角度,人們提出了共生起源學說、細胞分裂誤記說和DNA糾錯說等。有學者認為,在很多動植物中,“性”往往與休眠現象聯繫在一起,產生後壁孢子、種子或休眠卵,即休眠起源說,但分子機制有待進一步揭示 。
成活機率
雌性動物的卵子中只擁有母體一半的基因,如果卵細胞獨自發育成生命體的話,後代也只擁有母體一半的基因,它能成活嗎?答案是:能。一般人理解基因,會覺得它像一台鐘表一樣精密,增加減少一分都不可能。但其實基因中含有大量的無用信息,所以,有時候並不擁有和母體數量相同基因的後代也能正常發育。只擁有母體一半基因的個體叫“單倍體”。最典型的“單倍體”動物就是雄蟻,它是由蟻后的未受精卵發育成的。可以說,所有蟻后都能孤雌繁殖,而且要繁殖多少就繁殖多少,但有個問題:這樣生出來的都是雄蟻,而雄蟻是不能作為工蟻,也不能直接生下後代的。
如果要生出不能繁殖的雄蟻———工蟻或者可以繁殖的雌蟻———新蟻后,就需要產生出具有基因數目和母體相同的生殖細胞。當然,因為沒有和雄性交配,這些基因不可能從別的個體那裡獲得,只能母體“自力更生”。那么,母體又是如何自然產生二倍體(擁有和母體一樣數目的基因)的呢?
原來,在產生卵細胞之前,生物會先產生一種“卵原細胞”。它直接由體細胞分化而成,擁有和母體一樣數目的基因。卵原細胞經基因複製形成初級卵母細胞,初級卵母細胞第一次分裂,形成1個第一極體和1個次級卵母細胞。第一極體和次級卵母細胞再分別分裂形成3個第二極體和卵細胞,卵細胞和極體都擁有母體一半的基因。在正常情況下,缺乏營養的極體很知趣地消失了,擁有母體一半基因的卵子日後和來自雄性的精子結合,形成受精卵,從而發育出新個體。
如果在第二次減數分裂的時候,卵細胞與任意一個極體結合,形成“極體—卵細胞受精卵”,並由此細胞發育成後代個體,後代還是個“二倍體”。在這種情況下,實際上是把原本分配給卵細胞和極體的基因又摻和到一起去了,形成的極體—卵細胞受精卵的基因構成其實和卵原細胞———也就是母體的基因完全一樣。這樣產生的後代就是母體的“克隆體”。“亞馬孫螞蟻”可能就是這樣繁殖的。
還有另一種情況是,卵原細胞在進行減數第一次分裂時正常分裂,但不進行減數第二次分裂,最後形成了1個極體和1個“雙套卵”(diploid)。它攜帶有母體的全套遺傳物質,可以獨立地發育為後代個體(後代為二倍體)。這種情況雖然後代的基因數目和母體相同,但因為它是由彼此不同的卵子基因加倍形成的,所以兄弟姐妹之間彼此不同,而且後代和母親也不完全一樣。
最後一種情況是,卵原細胞正常進行了第二次減數分裂,產生3個極體和1個卵細胞,但任意兩個極體間發生了融合形成了“受精卵”。這也不是“克隆”。
簡單來說,孤雌生殖可能會產生單倍體,也可能會產生二倍體,這些二倍體可能是和母體完全相同的“克隆體”,也可能是彼此不同的“走樣複製品”。在科學家們對蛾子孤雌生殖的研究中,這些情況都曾遇見過。
特殊動物
青蛙的卵通過特殊的物理刺激也可孤雌生殖。
研究進展
1:在哺乳類動物中已實現人工孤雌生殖。
2:瑞士伯尼爾大學科學家伊爾邁澤於1981年成功地把小鼠體外受精卵的卵核和精核取出,植入雌鼠的未受精卵的卵核,繁殖出僅保留雌鼠遺傳基因的小鼠。
優劣
 孤雌生殖
孤雌生殖 有性生殖顯然大有好處,但是也有很多問題。
首先,有性生殖要耗費大量的能量。添加性交本身就已經比分裂生殖更麻煩了,更何況還要“尋找配偶”。如果同種生物的密度大還好說,像駝背鯨那樣幾百公里不見一頭的生物就麻煩了。所以座頭鯨一直“唱歌”以求知己。如果只是耗費能量還好,這個過程還很危險,因為不但伴侶能收到你的信號,捕食者也可以。無論是鳥鳴、蛙叫還是蟬噪,小動物們都懷著對雲雨之歡的嚮往一展歌喉,可招來的往往是野貓和獵隼。
尋找異性已經夠麻煩,跟同性競爭更耗費能量。無論是羚羊的鐵頭對攻還是松雞賽羽,都只為取得一個延續後代的機會。如果這種求偶競爭最終演化出累贅的身體器官,比如愛爾蘭麋過分臃腫的大角或者孔雀的長尾羽,那么這類生物本身都要受到被捕殺的威脅。
對於社會性生物來說,有性生殖嚴重威脅“安定團結”。人類中不斷發生的情感糾紛就不用說了,就連看起來“安分守己”的工蜂和工蟻都有可能為了自己能夠繁殖對女王造成威脅,揭竿而起。還有一點,大家可能沒有注意到,有性生殖產出的子女再親,也只有自己的一半基因,哪有“克隆”和自己那么相像?
大自然中的生物中存在著巨大的“退化驅動力”,簡單地說,只要環境允許,能簡單就簡單,能懶就懶。如果身處沒有地面天敵的環境,鳥類就會放棄飛行的能力。同樣,很多生境中的動物都會退回“不用交配就能生育”的狀態,也就是“孤雌生殖”。比如母蚜蟲,如果食物充足,它會抓緊時間進行孤雌生殖,能繁殖幾代就繁殖幾代,到秋末冬初時才會產生雌、雄兩性個體,兩者交配產卵越冬。
問題是,孤雌生殖又是怎么實現的呢?
科學家依然在探索。
