向性運功定義
植物的向光、向水、向地、向肥性、向重心性等運動方式的總稱。
tropic movement
植物體受到單一方向的外界刺激而引起的定向運動,稱為向性運動。
高等植物不能象動物一樣自由移動整體的位置,但植物體的器官在空間可以產生移動,以適應環境的變化,這就是植物的運動。高等植物的運動主要有兩種類型:向性運動和感性運動。植物向性運動是指在刺激方向和誘導所產生運動的方向之間有固定關係的運動。依外界因素的不同,向性運動主要包括向光性、向重力性、向觸性、向化性和向水性等。向性運動大多是生長性運動,是不可逆的運動過程。
1 向光性(phototropism)
植物隨光的方向而彎曲的能力稱為向光性,向光性是植物為捕獲更多光能而建立起來的對不良光照條件的適
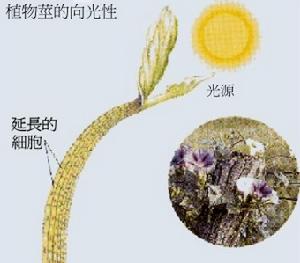 植物的向光性
植物的向光性藍光是引起植物向光性最有效的光源,研究表明植物體記憶體在著對藍光具有特殊敏感性而引起向光性反應的光受體。光生物學家已克隆出數種cDNA,並證明至少有1種對向光性受體負責,但至今未鑑別出何種cDNA表達向光性受體。紅光對誘導向光性反應無效,但有研究表明紅光對其有深刻影響。Liu和Lino(1996)證明紅光對藍光誘導向光性的影響是由光敏色素(phytochrome)介導的。在黑暗中生長的植物無向光性反應。該研究為生物學家找到光受體,闡明光轉導途徑提供了新的思路。
1.2 向光性運動機理目前,植物向光性運動機理有兩種假說:生長素分布不均勻假說和抑制物質分布不均勻假說。
1.2.1 生長素分布不均勻假說 Cholodny(1927)和Went(1928)以燕麥胚芽鞘為材料研究發現在單側藍光作用下,背光側胚芽鞘頂端擴散到瓊脂中的生長刺激物質活性高於向光側,並認為該物質是生長素,據此提出向光性運動是由於在單側光作用下生長素分布不均勻引起的,Thimann等(1937)稱之為Cholodny-Went學說,並套用該學說解釋植物向光性及向重力性運動現象,沿用至今,成為解釋向光性運動的經典理論。然而Bruinsum等(1975)以向日葵幼苗為材料研究植物向光性運動特性時發現,單側藍光處理並未引起生長素分布不均勻,因而對該假說提出質疑。Hasagawa(1989)重複Went試驗發現Went測得的胚芽鞘彎曲度是生長素和生長抑制物質兩類化合物總反應的結果,認為Went的結果是一種假象。但lino(1991)以玉米為材料,對玉米胚芽鞘向光側和背光側內源生長素含量的測定表明,單側藍光引起生長素分布不均勻,從而引起胚芽鞘向光彎曲,此結果支持了Cholodny-Went學說。
1.2.2 抑制物質分布不均勻假說 Hasagawa等(1980~1986)以蘿蔔等為材料,對向光性運動機理進行了詳細的研究,獲得了與Brunsum相同的結果,同時發現單側光引起向光側積累生長抑制物質,從而提出Brunsum-Hasagawa學說(1990),認為植物向光性運動是由於單側光引起生長抑制物質分布不均勻所致。一些研究表明,在不同植物中引起向光性運動的抑制物質有所不同。引起綠色向日葵幼苗下胚軸向光彎曲的抑制物質是黃質醛等(Franssen&Bruinsma,1981),引起蘿蔔下胚軸向光彎曲的抑制物質是順式蘿卜寧、反式蘿蔔寧和蘿蔔酚臉(Hasagawa等,1986)。燕麥胚芽鞘中的抑制物質至今未鑑定出,但根據色譜分析,它不是脫落酸。陳汝民(1999)研究指出抑制物質6-甲基2-苯並唑啉酮分布不均勻是單側光作用下玉米胚芽鞘向光運動的主要原因。
目前,在解釋植物向光性運動的兩種假說中,經典的Cholodny-Went學說面臨嚴峻的挑戰,似乎Bruinsum-Hasagawa學說占優勢。兩種學說尚有爭議,學者們還沒有統一的看法,尚待進一步深入的研究。
用鋁箔把光下生長的蒼耳葉片一半遮住後,葉柄相應的另一側延長,向光源方向彎曲,這樣葉片就會從蔭處移動到光亮處,葉片不易重疊,這就是葉鑲嵌現象。有人推測葉片遮蔽部分運輸較多生長素到該側的葉柄,因此該側葉柄生長快,葉柄向光彎曲。
1.4 太陽追蹤棉花、向日葵和花生等植物頂端在1 d中隨陽光而轉動,即所謂的太陽追蹤,葉片與光垂直,這種現象是溶質控制葉枕的運動細胞而引起的。向日葵的向陽運動是一個複雜的過程。葵花白天隨太陽由東而西轉動,正午時分朝南而轉向西方,到夜間八點半鐘左右由西而東轉,至於夜葵花已朝向東方而並非在第2天太陽初升時才從西而轉向東方的。葵花的運動機制尚無深入的研究。
2 向地性(gravitropism)
向地性是植物在重力影響下,保持一定方向生長的特性。目前,對向地性的研究已發展成為一新興學科一重力植物生理學。重力植物生理學在當代空間生命科學中具有舉足輕重的地位,肩負闡明地球重力在生物進化進程中的作用和空間不同重力環境中發展植物栽培技術的雙重任務。向重力性分為向地性、背地性和橫向重力性(地下莖水平方向生長)。
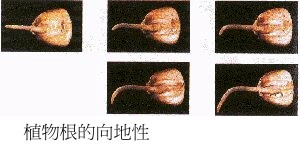 根的向性
根的向性目前,解釋根的向重力性運動機理有3種假說:Cholodny-Went學說、平衡石學說和雙叉理論。
2.1.1 平衡石學說 早在達爾文時代就已知道根冠是感受重力刺激的部分,但根冠如何感受重力刺激,長期以來眾說紛壇,其中Haberland-Nemee的平衡石學說受到普遍的重視。平衡石學說認為根冠感受重力刺激的是細胞內可移動的顆粒,稱之為平衡石(statolith)。一般認為,造粉體(amyloplast)可能是根冠細胞中感受重力作用的平衡石或稱之為重力感測器(gravisensor)。l個造粉體細胞含有 10~20個直徑為 4~10 um的澱粉粒。含平衡石的細胞稱為平衡細胞(statocytes)。植物體內的平衡石分布因器官而異。根部的平衡石在根冠中,而莖部的平衡石分布在維管束周圍的1~2層細胞(澱粉鞘)。平衡石在重力影響下,下沉在細胞的底部。
試驗表明,鈣離子在向重力性反應中起著重要的作用。將含有鈣離子螫合劑(如EGTA)的瓊脂塊放在橫放玉米根的根冠上,無向重力性反應;如改用含鈣離子的瓊脂塊,則恢復向重力性反應。進一步研究發現,玉米根冠中的鈣調素濃度是伸長區的4倍。外施鈣調素的抑制劑於根冠,則根喪失向重力性反應。
結合平衡石、生長素、鈣離子和鈣調素對向重力性的影響,有人提出向重力性的機理:根橫放時,平衡石沉降到細胞下側的內質網上,產生壓力,誘發內質網釋放鈣離子到細胞質內,鈣離子和鈣調素結合激活細胞下側的鈣泵和生長素泵,於是細胞下側積累較多的鈣離子和生長素,影響該側細胞的生長。
2.1.2 雙叉理論 Mesland(1992)根據非線形不平衡熱力學理論,提出重力整體作用概念,即所謂的雙叉理論。雙叉理論認為細胞核和細胞骨架在細胞對重力感受和傳導中起著很重要的作用,推測在細胞中可能存在重力致敏窗(gravity-sensitive window)。
Lorenzi&Perbal(1990)以扁豆幼苗為材料,通過不同處理條件下細胞核行為的研究發現,細胞核和細胞骨架確實起著重力感受器的作用。
劉承憲(2001)提出:重力感受的起始過程是根冠細胞中胞溶性Ca2+濃度局部增加,重力誘導胞溶性Ca2+濃度增加的機理,可能涉及到造粉體和細胞器的沉降,通過細胞骨架引起磷酸肌醇水解,胞溶性Ca2+濃度增加,活化鈣調蛋白和刺激Ca2+和Ca2+-鈣調蛋白依賴型的酶,如Ca2+-ATP酶和蛋白質激酶,最後導致胞內和胞外Ca2+梯度形成,不對稱Ca2+分布可能有差別的修飾細胞骨架蛋白,改變維管方位,影響細胞壁合成和沉積,結果形成一種非刺激側更快生長的生長梯度,產生彎曲。
禾穀類作物的莖有負向重力性反應。玉米和高粱節間的基部膨大,小麥、水稻和燕麥的葉鞘基部有特殊感受重力器官(假葉枕)。當這些植物的莖橫放或植株倒伏時,感受器官中的平衡石在2~10 min內便沉降到細胞下側,15~30 min內開始呈負向重力性反應,下側積累較多的生長素、赤黴素和乙烯,生長快,節間向上彎曲生長。
3 向觸性
達爾文在觀察西番蓮卷鬚向支柱快速彎曲運動時發現,卷鬚的末梢接觸到支柱後,在 20~30 S內就能激發出明顯的彎曲來。他認為這樣快速的運動不是向光性生長的機理所能解釋的,並大膽提出:卷鬚的向觸性運動是靠電波傳遞和原生質收縮來實現的。
經過以絲瓜卷須為材料的大量研究,婁成後等(1996)提出:卷鬚的快速向觸性運動是靠動作電波傳遞引起下段組織原生質體收縮來完成的,動作電波的傳遞也不是單靠局部迴路電流,還要有神經遞質乙酸膽鹼的相互協作,交替推進來執行。
此外,植物的向性運動還有向化性和向水性。向化性是指某些化學物質在植物周圍分布不均勻而引起的生長,如作物根部朝向肥料較多的土壤生長。
向水性是指土壤中水分分布不均勻時,植物根趨向較濕地方生長的特性。
在單側光的照射下,植物朝向光源方向生長的現象叫做向光性。
