
學說介紹
中性學說(the neutral theory)認為分子水平上的大多數突變是中性或近中性的,自然選擇對它們不起作用,這些突變全靠一代又一代的隨機漂變而被保存或趨於消失,從而形成分子水平上的進化性變化或種內變異。
 中性學說圖解
中性學說圖解1968 年,日本遺傳學家木村資生(1924--1995)根據分子生物學的研究資料,首先提出了分子進化的中性學說(The neutral theory of molecular evolution),簡稱“中性學說”(the neutral theory)或“中性突變的隨機漂變理論”。1969年美國學者J·L·金和T·H·朱克斯又用大量分子生物學的資料肯定了這一學說。
提出背景
20世紀50年代以來,科學家先後搞清楚了許多生物大分子的一級結構。通過比較不同生物的某些功能相同的蛋白質的胺基酸序列或核酸的核苷酸序列的差異,人們發現,親緣關係近的差異較小,親緣關係遠的差異較大,與物種的表型進化情況基本一致。分子進化至少有三個顯而易見的特點:一是多樣性程度高,與表型多態(即在一相互交配的群體中存在著兩種或多種基因型的現象)相比,分子多態更為豐富(例如細胞色素C這種蛋白質分子在行有氧呼吸的不同物種中就有種種不同的分子結構);二是各種同源分子對選擇大都是中性或近中性的,它們都有完整的高級結構,能很好地完成各自的功能(如脊椎動物的血紅蛋白分子都能運氧、各種生物的細胞色素C都能在氧化磷酸化中完成電子的傳遞等);三是隨著生物從低級向高級演化,同源分子中逐年發生胺基酸或核苷酸的替換,且大致按每年每位置替換數恆定速率進化,也就是說,每一種生物大分子不論在哪個生物體內,都以一定的速率進化著。例如,各種脊椎動物血紅蛋白的分子α鏈中的胺基酸,是以每年大約10的負九次方個的速率置換著,並且置換的速率與環境的變化和生物世代的長短無關。關於大分子的進化性變化,早在1965年E·楚克坎德爾和L·C·波林以及 E·馬戈利亞什和G·H·史密斯就已有過全面的論述。木村的功績是在理論上更邁進一步,把中性突變-遺傳隨機漂變放到決定性的位置上,提出分子進化的中性學說,較合理地解釋了分子進化的各種現象。
學說依據
 相關圖示
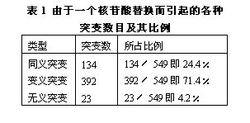
相關圖示中性學說的出發點是中性突變。DNA(脫氧核糖核酸)產生突變的方式有四種,即核苷酸的替換、缺失、附加和倒位,而最常發生的是核苷酸的替換。對一個密碼子來說,由於核苷酸替換可發生在三聯體的任一位置上,而每一位置都可能有三種替換方式,故一個核苷酸替換可有九種方式導緻密碼子變換,如AAA→AA(C、G、U)AAA→A(C、G、U)A AAA→(C、G、U)AA。因此,61種胺基酸密碼就可因一個核苷酸替換而產生(61×9=)549種密碼子變換。AAA與AAG都是Lys的密碼子,這種密碼子變換並不引起胺基酸替換,稱同義突變或沉默突變。AGA是Arg的密碼子,UAA是終止密碼子,從AAA到AGA引起胺基酸替換,稱為變義突變;從AAA到UAA 則稱無義突變,因為UAA代表終止信號。如果將549種變換對照遺傳密碼錶逐一統計,可以發現由於遺傳密碼的簡併就保證了約1/4的替換為同義突變(表1)。
 相關圖示
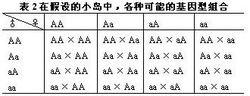
相關圖示胺基酸替換又分為不影響蛋白質功能的、性質相似的胺基酸之間的替換即保守替換(如Arg和Lys間,Phe和Tyr間,Glu和Asp間等)和影響蛋白質功能的、性質明顯不同的胺基酸之間的替換即非保守替換(如 Asp和 Val間,Phe和Ser間,Arg和Ile間等)。若每種方式以同樣機率發生,則由一個核苷酸替換所引起的全部胺基酸替換中有53%是保守替換,而從分子進化研究中觀察到的胺基酸替換來分析,保守替換可高達68%。同義突變和保守替換不改變或很少改變分子的結構與功能,所以在選擇上是中性的或近中性的。如上所述,這類變異在生物體內大量存在。此外,DNA的非編碼區以及基因的內含子區所發生的核苷酸替換,以及不轉譯成蛋白質的假基因中所發生的變化(這些都是分子進化研究的對象)等,也是中性或近中性的。自然選擇對中性或近中性的突變不起作用,在眾多的中性突變中,哪一種變異能夠流傳,哪一種變異趨於消失,全靠機遇,這種基因頻率的隨機變動就是遺傳漂變。遺傳漂變最初由S·賴特於20世紀30年代提出。他認為,當一個大群體劃分為若干個小群體繁殖時,群體的基因組成實際上等於進行了一次隨機取樣。例如,假設有16個小島,每個島上都有遺傳型為Aa的公羊和母羊各一頭。這一對羊相互交配,子一代的基因型應為1AA︰2Aa︰1aa。如果群體較小,每島僅留下雌雄羊各一頭進行繁殖,就會產生如表2所示的結果。即16個小島中,1個小島只留下AA雌和AA雄,A基因的頻率就從0.5變為1;另1小島只留下aa雌和aa雄,a基因就得到了固定;其他14個小島上,A和a則以不同的比例保存(表2)。而對16個島進行統計分析,基因A和a的頻率仍為0.5,三種基因型之比仍為1︰2︰1。由此賴特認為,遺傳漂變僅發生在小群體中,它對生物進化的貢獻是極其有限的。木村資生則證明,遺傳漂變並不限於小群體,對任何一個大小一定的群體,都能通過遺傳漂變引起基因的固定,從而導致發生進化性變化。他還提出,遺傳漂變是分子進化的基本動力。
主要內容
突變大多是“中性”的
突變大多是“中性”的,它不影響核酸和蛋白質的功能,對生物個體的生存既無害處,也無好處。這類突變有“同義”突變、“非功能性”DNA順序中發生的突變以及結構基因中的一些突變。例如,在三聯密碼中,一個核苷酸發生置換,往往不會造成胺基酸的改變。如 UUU和 UUC都是苯丙氨酸的密碼子, C和U之間相互置換,不改變密碼子的功能,這兩個密碼子好比是“同義詞”,因此“同義突變”是中性的。木村資生曾把決定組蛋白Ⅳ的胺基酸排列的兩組海膽的mRNA進行某些比較,發現其中五個地方有變化,但胺基酸則完全一樣(表15-2)。鹼基變化了,胺基酸不變,就可以認為是中性。木村說,不改變胺基酸的鹼基置換是這樣進行的,“基因DNA的信息被mRNA所轉錄,其信息使鹼基進行排列,每三個鹼基決定一個胺基酸。它的第三個位置的鹼基被置換了,這個變化是同義的。就好比回答一個問題時說‘是’。找出可以把‘是’字換成‘對’字。雖然字改了,但意義相同。像這樣第三個位置上的鹼基置換,一般是可以的”,不僅如此,而且在上述的海膽中這種不改變胺基酸的鹼基置換進行的速度非常快。金和朱克斯還認為,這樣的突變頻率,約占核苷酸置換的1/4。
隨機固定
“中性突變”通過隨機的“遺傳漂變”在群體裡固定下來,在分子水平進化上自然選擇不起作用。中性學說認為,當一個生物體的DNA分子出現中性突變,既不提高也不降低它在生活環境中的生存適合度,它是通過群體中的隨機交配,使這些突變在群體裡得到固定、發展或者消失。這一學說認為,“遺傳漂變”在進化中起重要作用是有據可循的。例如,許多不同物種的功能相同的蛋白質,如血紅蛋白、細胞色素c、核酸酶、胰島素、免疫球蛋白,血纖維蛋白肽等,它們的胺基酸組成是有很大區別的。又如,兩種可以交配並產生子代的蛙(Xencpus leavis和Xenopus mulleri),它們的DNA中的一些重複順序的相差程度可達10倍到100倍之多。這一學說認為,上述情況說明,不受自然選擇壓力的中性突變,通過隨機的“遺傳漂變”在群體中得到固定和逐漸積累,可以實現種群的分化,出現新的物種。
中性突變決定進化的速率
第三,進化的速率由中性突變的速率所決定,也就是由核苷酸和胺基酸的置換率所決定。它對於所有的生物,幾乎都是恆定的。木村資生認為,在表現型水平的進化中,進化速度也是非常快的,也有像所謂“活化石”那樣進化極慢的類型。但是,基因水平上進化速度幾乎是一定的。中性說最初的論文,首先就是論述進化速度問題。
這一學說認為,如在體內運載氧的紅細胞中的血紅蛋白β鏈由 141個胺基酸組成,各個胺基酸每年以平均 10-9(十億分之一)的比率變化著。就是說,對每個胺基酸來說,要10億年變化一次。在細胞內擔任氧化還原的細胞色素c,以血紅蛋白的1/3的速度在變化。血液凝固的時候,從血纖蛋白原脫變成血纖蛋白時所放出的血纖蛋白肽,比血紅蛋白的進化速度快幾倍。由此可見,分子種類不同,分子的置換率不同,進化的速度也不同。但是,同一分子的進化速度在不同物種中卻是相同的。而且與世代的時間長短無關。
層次
根據金和朱克斯的計算,決定胺基酸的密碼子中鹼基替換的速率為每年每個密碼子替換3-50×10-10個。根據恆定的置換速率和不同物種之間同種蛋白質分子的差射,可以估計出物種進化的時間。這一學說認為,這種計算的結果,同根據化石研究所確定的進化時間十分接近。由此可見,達爾文主義、現代達爾文主義和中性學說在生物進化問題上研究的層次是不同的。前者是從個體或群體的層次出發,後者則是從分子水平的層次出發。兩者之間的主要區別是:前者認為生物進化的主導因素是自然選擇,後者則認為生物進化的主導因素不是自然選擇,而是中性突變本身;前者認為生物進化的方向與環境有必然的聯繫,後者則認為生物進化的方向與環境無關,純粹是生物分子隨機的自由組合;前者認為生物進化的速率受環境和生物世代等因素的制約,後者則把生物進化速率的一致性、恆定性作為分子進化的主要特徵。由於中性突變不受自然選擇的作用,中性突變的速率就等於分子進化的速率。
中性說是從分子水平研究上提出的一種新的進化學說。這一學說在學術界引起較大的反響。由於分子生物學的發展,基本上弄清了分子以及基因之類的內部結構,也能在數量上對此進行控制,因此木村對分子或基因的進化就能依靠數學的理論來進行處理。這樣,自然選擇不能解釋的那種出於意外地進化姿態也就出現了好幾種中性突變的類型。同時,中性說套用了分子生物學的技術和數學方法,打破了不同物種之間不能進行雜交試驗的局限性,可以對不同物種的同源蛋白質胺基酸順序和基因進行比較,並計算出分子進化的過程和速度。這樣,進化論的研究中定量的水平便更高了。只要有了突變率、遷移率和群體大小等參數,還可以預測任何一個特定群體中遺傳變異的數量和基因頻率變化的速率。此外,中性說學者還認為,即使在表現型水平上,也不能完全否定中性說的作用,不能完全肯定達爾文學說的正確。正如木村所說。“……假如分子水平中性說是正確學說的話,那么表現型水平就也有中性了。說達爾文自然選擇是百分之百的正確也許不切合實際吧。例如,對人類來說,雖然有鼻子高或鼻子低的,但對其生存來說究竟有多少利弊呢?真正的自然選擇作用是有餘地的。我認為這樣的差異,對於生存是既不好也不壞的”。
進化速率
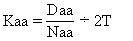 式1
式1分子進化速率是以每年每位置胺基酸或核苷酸替換數來表示的,以蛋白質分子為例,其進化速率如式1:
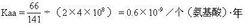 式2
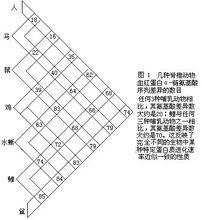
式2式中,Daa是兩種不同生物同源蛋白質的胺基酸差異數,Naa是構成同源蛋白質的胺基酸個數,T是兩種生物的分歧進化時間。公式一中之所以要除以2T,是因為分歧進化是向兩條路線進行的。可以這樣理解:從一種生物回溯到分歧點再到第二種生物,其演化時間正好是2T。分子進化的特點之一是每一種生物大分子不論在何種生物都有一個大致恆定的進化速率。以血紅蛋白α-鏈為例,鯉與馬有66個胺基酸差異(圖1),地質資料表明魚類約起源於4億多年前的志留紀,若以4億年作為魚與馬的分歧進化時間,則從魚到馬的進化速率如式2:
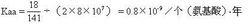 式3
式3馬與人有18個胺基酸差異(圖1),高等哺乳動物的輻射進化約出現於8 000萬年前,如以此作為馬與人的分歧進化時間,則從馬到人的進化速率為:
即血紅蛋白的α鏈分子,無論是在從魚到馬還是在從馬到人的進化過程中,其進化速率基本上都是相同的。分子進化速率與種群大小、世代壽命和物種的生殖力均無關,也不受環境因素所影響。這是自然選擇學說所不能解釋的,而中性學說對此卻能作出既簡明又合理的解釋。
設大小為N的一個二倍體群體中,共有2N個基因位點,若每配子每代的突變率為υ,則每代將產生2Nυ個新突變。設一個突變最終被固定的機率為μ,則每代每位點的突變替換數(進化速率)為:K=2Nυμ
爭議
邁爾(2009)對木村的理論進行了猛烈的抨擊:考慮到基因並不是自然選擇的靶子,因此,所謂的中性進化就是一個毫無意義的概念 。
謝平(2014)認為,分子鐘或中性理論的倡導者與達爾文一樣都關注隨機變異(當然前者在分子水平,後者在表型水平),只不過前者對不受自然選擇作用的那些變異感興趣,而後者則關注受到自然選擇作用的那些變異;可是,任何水平的變異既具有隨機性,又不可能完全隨機,而分子鐘或中性理論的信奉者就是要找出既普遍存在又逃脫了達爾文選擇的神奇分子,或者說一個普適性存在且完全不受達爾文選擇影響的分子就是他們夢幻中的時鐘分子。當然,這樣的分子還必須具有足夠的複雜性,以便容納相當大的時序性變異,而過於簡單的分子是不可能作為這種時鐘分子的 。
細胞色素C是一類對中性理論起到了關鍵作用的分子,它既複雜、又廣泛分布(作為電子載體存在於幾乎所有的生物類群之中)且變異也十分緩慢,看上去十分符合理想時鐘分子的條件。但是,雅荷雅(2003) 指出,一些研究表明,這種分子在生物中表現出難以置信的變異:兩種不同爬蟲間的差異,要比鳥跟魚或魚跟哺乳動物的大,同一鳥類分子間的差異,比這類鳥與哺乳動物的還要大,此外,看上去相似的細菌,在分子中的差異,要比哺乳動物、兩棲動物或昆蟲的大。類似的例子應該舉不勝數!
謝平(2013)指出,由於蛋白質決定生物有機體的結構和功能,因此,它應該是表型變異的分子基礎,如果自然選擇作用於表型,就必然作用於控制表型的物質基礎—蛋白質。因此,從邏輯上來說,分子與表型、個體乃至物種之間的進化關係不可割裂 。
Denton(1985)指出,“在分子水平上的每一生物種類是獨特、孤立的,並且與任何媒介沒有關聯。因此,像化石一樣,分子沒有給長期尋找難懂的中間形式的進化論生物學家以證據……。在分子水平上,任何生物體沒有可以比作‘祖先’、‘原始’或‘高級’的親緣關係……。無疑,如果這些分子證據在一個世紀以前存在的話……器官進化的觀念也許根本不會存在” 。
