
現象發現
光周期現象是美國的Garner和Allard發現的。上述現象說明植物在特定季節開花,他們認為一定有某個環境因子在控制開花,大家知道,主要的環境因子有溫、光、水、氣、礦質營養,那么隨季節變化的主要是溫度和光照長度,因此,他們檢驗了日照長度對菸草開花的影響,結果發現,只有當日照短於14小時時,菸草才開花,否則就不開花。後來又發現許多植物開花需要一定的日照長度,如冬小麥、菠菜、蘿蔔、豌豆、天仙子等,這就是光周期現象的發現。1920年,Garner和Allard在美國馬里蘭州,美國農業部Beltsville農業試驗站工作,他們發現兩個難以解釋的現象,一個是菸草品種馬里蘭猛獁象,在夏季株高可達3~5米,但是不開花,如果在冬季的溫室里,株高不到1米就可以開花;另一個現象是,某個大豆品種,在春季的不同時間進行播種,但在夏季的同一時間開花,儘管不同播種期大豆的營養體大小不同。生物反應
生物對晝夜光暗循環格局的反應。光周期指晝夜周期中的光照期。20世紀20年代植物生理學家首先對光周期現象進行了科學研究,當時認為主要是光照期的長度決定植物是否進入開花期。但後來的研究表明,暗期的長度對開花極為重要。事實上光照期和暗期的相對重要性因物種而異,有時兩者都不可缺。每日光照期的長短因季節和緯度而異:如在北半球冬季日照短而夏季日照長,在南半球則適相反。季節間日照長度的差異在赤道附近不明顯,但從赤道越往高緯度越顯著。對一定的緯度來說,日照期的長度可以準確地指示季節,比日照強度和氣溫都更可靠。在北半球的秋季,隨著日照時間的縮短,植物開始落葉,某些昆蟲進入滯育,候鳥準備南飛;至春季隨著日照時間的延長,候鳥北返,昆蟲復甦,植物也紛紛進入著葉開花期。在這些現象中都是日照長度的變化給生物提供了信息。光周期現象是物種長期進化的產物,它保證了生物能在適宜的環境條件下生長和繁育。生物光周期現象因生物本身的發育階段而異,同時也受氣溫、食物等外界環境因素的影響。現已知,光周期現象是和生物內在的生物鐘機制耦合在一起而發揮作用的。
人類早已將有關光周期現象的知識用於生產,套用較廣的實例如調節光照時間以控制花卉的開花時間和家雞的產卵量。在自然條件下,晝夜周期的總長度是個定值(24小時),一定的光照期必然對應一定的暗期。在人工條件下,雖可以產生任意長度的光暗組合,但要產生預期效果,仍需大致模擬生物已長期適應了的自然情況。
植物、昆蟲和鳥類的光周期現象比較明顯,對它們的研究也比較多。這包括植物的開花、落葉,種子和芽孢的休眠及塊莖、塊根的形成等;昆蟲的滯育、遷徙和型變等;鳥類的遷飛、生殖腺發育、體脂積累和換羽等。生物必需有感光機制才能對光周期作出反應:在植物中有光敏素;在動物,視神經以及松果體或其他腦部結構是反應的第一站。其次,生物體內必需存在測時裝置(生物鐘)才能判定光照期(或暗期)的長度是否適宜,這方面的研究還沒有取得什麼重要成果。最後,在反應的傳出環節上,植物可能是通過激素;動物則包含神經和激素兩者。生物光周期現象是進化形成的遺傳特性,它決定了物種能對光周期作出什麼反應,但實際發生的反應卻又受環境條件的影響。對親蠶進行光照實驗,反應卻表現在子代上;光照長短可決定子代是否滯育。這都說明光周期現象的複雜性,需要結合生理學、遺傳學、生態學等多方面的研究才能闡明。
植物
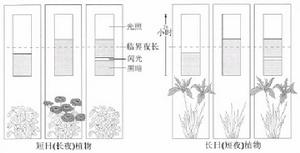 植物的光周期現象
植物的光周期現象在一些植物葉部存在的光敏素,可能是有關的感光物質。在實驗中,一個植株接受適宜光照,與之嫁接的另一未受光照的植株也發生開花反應,這說明是經激素傳導,不過至今尚未分離出有關激素。
鳥類燈草鵐在春季的生殖腺發育和向北遷徙是光照期變長的結果,如果在晚秋和冬季人工加長光照時間,也可誘導其生殖腺早熟和提前北飛。現已知道多種鳥如燕雀、鵐、麻雀、掠鳥等都有光周期反應,主要表現在生殖腺發育,脂肪體積聚、脫換羽毛和遷徙等方面。
 鳥類光周期現象
鳥類光周期現象春季鳥類的遷徙受光周期控制,這是溫帶和亞熱帶許多旅鳥的特徵。在熱帶這種現象並不明顯。在溫帶許多於南方過冬的鳥,如鵐、掠鳥等,經歷冬季和早春的短日照階段,生理上發生一系列變化,這一階段稱為“生理準備期”。隨著春季光照時間的日漸延長,鳥體內逐漸積累大量脂肪體,生殖腺逐漸增大。當生理變化達到一定階段,且光照時間達臨界值以上時,鳥就開始向北遷飛,到達繁殖地區。這種變化隨光照時間的增長而加快。秋季的短日照階段對鳥類來說是必要的,稱為生理反應的“準備前期”。未經歷此期的鳥不能在隨後的冬季和早春實現一系列生理反應過程。
鳥類的光周期反應主要表現在光照期。這與植物迥然不同。光照的強度對鳥類生殖腺的光周期反應並無明顯影響,只需達到某一最低光照強度即可。每種鳥也都有自己的最低感光值,但增加光的強度並不能加速光周期反應,重要的仍然是光照時間的長短。光照期的作用光譜一般是橙黃色至紅色的光,更傾向於紅光(600~750納米)。
有些鳥類的更換羽毛也受光周期控制。長光照可誘導白喉帶鵐、白冠帶鵐等換毛,人工延長每日的光照時間往往可促其提前更換羽毛。
昆蟲多種昆蟲的滯育、遷徙及型的變化都受光周期的控制。許多昆蟲可以在嚴寒、酷暑來臨之前,逐漸降低新陳代謝、停止生長和發育,以順利渡過這極端時期。這稱為滯育,而滯育的誘導主要靠光周期的變化。在春季的短日照條件下家蠶產不滯育的卵,但在24℃飼育幼蟲並每天光照16小時以上,則幼蟲發育成蛾後所產的卵全是滯育卵。因此,家蠶可稱為短日照型昆蟲。
昆蟲一般是在發育早期受光周期的誘導,至發育後期才表現出滯育。玉米螟幼蟲對光周期的感應是在3~4齡期,滯育出現在前蛹期。番茄夜蛾的幼蟲在8小時短光照條件下飼養,至蛹期才滯育。
許多昆蟲的遷徙與光周期有密切關係。在北美有一種大紅斑蝶,夏季在加拿大和美國北部生長繁殖,秋季日照縮短後就成群結隊向南遷移至墨西哥過冬,翌年春季日照增長後又北遷。短光照促使馬鈴薯葉甲從寄主植物轉移入土壤休眠,也使七星瓢蟲遷移上山越冬。
蚜蟲在不同季節定居在不同種類的植物(木本或草本)上。無翅的個體還要轉化為有翅類型才能在不同生境間遷移。而且蚜蟲大部分時間營孤雌生殖,但入冬前卻要分化出兩性個體,進行有性生殖,再以受精卵越冬。這些型變也是受光周期誘導的。
昆蟲的光周期反應主要是對光照期長短的反應,但在短日照型昆蟲,光照與黑暗的交替似乎是主要的。短日照型昆蟲的光周期反應與植物不同,若在暗期中間給予短時間的光照,藉此把暗期分成數段,而暗期總的時間不變,在這種情況下昆蟲的光周期反應並不發生變化。這說明暗期不一定要連續,光照期卻可以累加。昆蟲光周期反應的另一個特點是它的光強度閾值低,即弱光就足以引起反應。昆蟲的光周期反應對光波性質有一定的選擇,最敏感的光一般是藍紫光(350~510納米),而黃色至紅色的長波段光無效應。
地理緯度
我國地處北半球,春季的短日時期氣溫較低,植物一般處於苗期,與開花暫時無關。秋季則氣溫較高,適於植物生長,所以這時的日照就顯著的影響植物的開花。夏季的自然長日照時期,氣溫較高,是植物生長發育的適宜時期。一年之中,影響植物開花結實的基本上是這兩個季節。在低緯度地區,終年氣溫較高,但無長日條件,所以只有短日植物,一般都是在早春出芽,在夏季和秋季任何時候均開花。有些植物可以多季,如水稻;在中緯度地區,既有長日條件,又有短日條件,且秋季氣溫較高,所以長短日植物均有分布。長日植物在春末夏初開花,而短日植物在秋季開花;在高緯度地區(我國東北),雖然有長日和短日條件,但氣溫的季節性變化比較明顯。秋季短日照時,氣溫已低,植物又不能生長。所以不能生存一些要求日照較長的植物。植物光周期反應的不同類型是長期適應環境的結果。由於地球上同一緯度在不同的季節、不同緯度在同一季節之間光周期不同,所以就形成了植物光周期反應類型的規律性分布。
