介紹
現已描述的種類約為175萬種,其中,昆蟲和多足類多達96.3萬種,植物約有27萬種,脊椎動物有5萬多種,古生菌只有175種。不同類別物種的分化速率有著天壤之別:為何昆蟲種類如此之多,而大象或長頸鹿種類如此之少?
分化速率
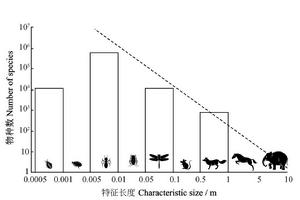 以特徵長度來分類的所有陸生動物物種數
以特徵長度來分類的所有陸生動物物種數May(1988)通過分析陸生動物物種數與體積之間的關係發現,在大部分體積區間,物種數與體積成負相關,也就是說,隨著物種體積的增大,新物種的分化越來越困難。一般來說,體積較小的動物在進化上較 為低等和原始,結構也相對簡單。達爾文曾指出,“在自然系統中低級的生物比高級的生物容易變異,是和博物學者的共同意見一致的。我這裡所謂低等的意思是指體制的若干部分很少專業化,以擔任一些特殊機能;當同一器官勢必擔任多種工作時,我們大概能理解為什麼容易變異,因為自然選擇對於這種器官形態上的偏差,無論保存和排斥,都比較寬鬆,不像對於專營一種功能的部分那樣嚴格”(Darwin 1872)。
為何動物物種的分化速率與體積成負相關?有幾種可能的原因:①動物的體積越大,繁殖速率越慢,因此,物種分化也就越慢;②動物的體積越大,內部機構就越複雜,形態與生理變異的可塑性相對變窄,因此新種的分化就越困難,③在一定的空間範圍內,動物的體積越大,所能容納的物種數就越少。一般來說,動物的活動範圍隨體重的增加而增加,而且同樣的體重,肉食性動物的活動範圍比牧食性動物更大,恆溫動物的活動範圍比冷血爬行動物要大。
繁殖速率(它也取決於體積大小或體制的複雜性)也許最為關鍵。對有性生殖的物種(幾乎所有動物都能進行有性生殖)來說,減數分裂過程中染色體的聯會與交換是變異的重要來源,因此,就不難理解為何繁殖速率越快,物種分化也快的現象了。
在絕大多數情況下(體長<1 cm的物種除外),r生態對策者似乎是物種快速分化的天堂,而極端K-生態對策者(如大象)幾乎就是進化的死角,它們似乎容易重蹈愛爾蘭麋鹿的滅絕之路。
組合物種
細菌和古菌合計起來也不過1萬種左右,這不足現存物種的1/170,巧合的是,在動物(真核生物)中,當體長小於1cm時,物種數也出現顯著下降。為何細菌的種類是如此的稀少呢?它們不是繁殖速率很快的r-生態對策者嗎?細菌種類的稀少有三種客觀的解釋:其一,微生物定種困難,因為絕大多數細菌無法進行純培養;其二,如果結構過於簡單的話,形態特徵的鑑定和區分變得越來越困難,容易被分類學家所忽視。但是,這些並不會從根本上動搖細菌種類稀少的事實。其三,當生命的結構簡化到極限的程度時,可能無法允許更多的物種存在。
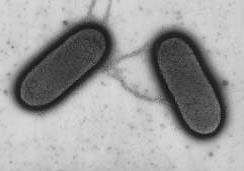 細菌之間的遺傳交換
細菌之間的遺傳交換雖然細菌種類不多,但是它們卻能夠通過快速的變異應對幾乎無限多樣的生存環境。這種快速的變異通過快 速的繁殖及個體間基因的重組得以實現—細菌的抗藥性快得驚人,根本不需要地史的時間尺度,此外,一些細菌之間能靈活地進行遺傳交換,這雖然在方式上有別於有性生殖物種在減數分裂過程中通過同源染色體之間的聯會與交換實現的頻繁的基因交換和重組。
不少證據表明,細菌的基因庫似乎具有驚人的變異性與流動性。赫奇斯甚至大膽地認為,“可以把基因看作一個巨大的公共基因庫,當細菌為了改變境況,即為了改變與變化中的環境的關係時,臨時定義的‘種’就從基因庫中獲取它們所需的遺傳信息”(詹奇1992)。
謝平(2014)指出,細菌之間都能快速而靈活地進行遺傳交換的事實似乎昭示著整個細菌界是由若干個超級物種所構成,因為在自養細菌(如藍細菌)與異養細菌(如病原菌)之間可能還是應該存在若干難以融合的根本性差異的。而真核生物則採取了完全不同的方式,即真核生物的一個物種就是一個相對獨立的基因庫,它們分化出了數以百萬計的物種基因庫。日益複雜化的真核生物顯然不可能再採用原核生物的這種臨時性物種組合遊戲了,因為複雜性(多細胞化、組織與器官分化等)不可避免地犧牲了簡單靈活性。總之,細菌的生存策略似乎是:以小的細胞體積、基因組的簡潔性、靈活的基因交流以及快速的無性繁殖方式來實現對無限多變環境的完美的適應性。
成種速率
一般來說,體積越大的動物體制也越複雜,這裡體制指生物的內部結構、組織形式和生理功能等,它們可用一些相關的參數來表征。謝平(2014)認為,越小的生物(如昆蟲)其個別基因的突變對體制突然改變的可能越大,而越大的生物其個別基因的突變對體制的突然改變的可能性越小,像對大象、人這樣極端複雜的生物這幾乎是不可能的。越小的生物,越能承受體制參數的大幅波動或損傷,或者說能通過快速適應固定發生變化的新體制,這樣可塑性也就越強。因此,生物體制的可塑性就意味著物種分化的潛力,可塑性越強,遺傳重建速度越快,物種就越豐富,譬如,小小的昆蟲有70-80萬種之多,而龐然大物—大象在全世界僅有3種!
表1 現在已知的生物主要類群的種類,包括尚未被發現的種類的估算
| 域 | 真核界 | 已描述的種類數 | 估計的總數 |
| 古生菌 | 175 | ||
| 細菌 | 10,000 | ||
| 真核生物 | |||
| 動物界 | 1,320,000 | ||
| 脊椎動物門(總計) | 52,500 | 55,000 | |
| 哺乳動物 | 4,600 | ||
| 鳥類 | 9,750 | ||
| 爬行動物 | 8,002 | ||
| 兩棲動物 | 4,950 | ||
| 魚類 | 25,000 | ||
| 有鄂動物(昆蟲和多足類) | 963,000 | 8,000,000 | |
| 螯肢動物(蜘蛛等) | 75,000 | 750,000 | |
| 軟體動物 | 70,000 | 200,000 | |
| 甲殼動物 | 40,000 | 150,000 | |
| 線蟲 | 25,000 | 400,000 | |
| 真菌 | 72,000 | 1,500,000 | |
| 植物界 | 270,000 | 320,000 | |
| 原生生物 | 80,000 | 600,000 | |
| 總計 | 1,750,000 | 14,000,000 |
(引自Groombridge and Jenkins 2002)
從生態對策來看,小的物種稱為r—對策者,譬如,相對於老虎(K-對策者)來說,蒼蠅和老鼠就是r-對策者,後者的生態可塑性也強。蒼蠅和老鼠能忍受種群的劇烈波動,可憑藉其驚人的繁殖率從極低的種群密度快速恢復,它們除之不盡;而老虎卻不能忍受種群的劇烈波動,十分容易被人類滅絕。顯然,體制的可塑性與這種生態對策相吻合,即r—對策者的體制可塑性也較強。或者說,生物的體積(或複雜性)與體制的可塑性呈負相關。
為何結構的複雜性會犧牲體制的可塑性呢?這既有遺傳的原因也有生態的原因。從遺傳本質上來看,有性生殖通過基因不斷堆積來創造變異與適應(最終新物種)的方式,不可避免地會導致基因組、表型、結構與調控的過度複雜化(在一些情況下還有體型的過度大型化),這就埋下了致命的隱患—可塑性的降低。從生態學來看,結構複雜的動物個體對中度以下的環境波動具有較好的緩衝性,但是,這卻犧牲了繁殖速率,也犧牲了更廣泛的變異能力與適應範圍。因此,一旦遭遇到環境災(巨)變,複雜的生命往往難以進行大幅度的改變與適應,不可避免地遭受滅頂之災,龐然大物恐龍的滅絕就是一個活生生的悲慘案例(謝平2013)。
基因及其調控還有生命整體絕不可能趨於無限複雜,終究會將發展途逕自我阻斷,使某個方向的進化嘎然而止。生命特有的不斷複雜化的遺傳進化方式可能是導致許多大型動物(如恐龍、愛爾蘭麋鹿)滅絕的內在機制之一。這種現象常常被直生論者稱為“內部完美原則”或內在的“種系動力”,最初是為了適應,但是由於這種適應的創造方式使機體越來越複雜,逐步降低了機體的可塑性(這與生態系統的複雜性與可塑性的關係完全不同),最終在環境巨變來臨時慘遭滅絕(謝平2014)。神創論者常常藉此來攻擊達爾文的變異。雅荷雅(2003)說,“變異不會通過給爬行動物增加翅膀或羽毛,或者改變其新陳代謝的方式,使它們轉變成鳥類”。
謝平(2013)指出,類似於生態(或生殖)對策,似乎也可以將成種區分為兩種不同的類型。較小的動植物(如昆蟲)繁殖速率快,體制的可塑性強,演化的方向多,突變成種的速率快,物種十分豐富,可稱之為r—成種策略(r-speciation strategy);而較大型動物(如哺乳動物)繁殖速率慢,體制的可塑性差,演化的方向少,突變成種的速率慢,物種數較少,可稱之為K-成種策略(r-speciation strategy)。至於微小變異的連續累積和用進廢退的獲得性遺傳,不論動物大小都會自然發生,都會貢獻於物種分化。總而言之,r—成種策略是快速的隨機性演化的天堂,而緩慢的K-成種策略往往會奔向直生論的死角。
當然,具有相似體制的物種之間偶爾也會在成種速率上出現巨大的差異。有一些物種很難發生成種事件(或成種速率很慢),被稱為“活化石”,而相關物種卻發生了劇烈的變化或者已經絕滅。譬如,在北美東部有一些植物(包括臭菘),也存在於東亞的一些地區,這些廣泛分布於兩個大陸的群體不僅形態上沒有什麼區別,而且雖然已經分離了600萬至800萬年,還可以相互交配。有些動植物物種經歷1億多年也沒有發生明顯變化,如動物中的鱉、仙蝦和酸漿貝,在植物中長期存在的屬有銀杏、南洋杉、木賊和蘇鐵(邁爾2009)。
有數據表明,物種的壽命分布是冪律的,大多數物種形成之後不久便滅絕了,只有少數物種才能存活較長的時間,這種物種形成與滅絕事件的冪律分布大約在6.5億年前的顯生宙時期便出現了(考夫曼2004)。因此,“活化石”物種似乎可用物種壽命的這種冪律分布予以解釋,雖然這還只是一種統計性的解釋。
