
資源競爭理論
資源競爭理論主要是以Monod 方程和Droop 方程為基礎而發展起來的。
Monod 競爭模型
Tilman將Monod 方程推廣至多種群間對於多種資源(如營養鹽) 的競爭。在多種群需要的多種資源中,對於n 個種群中的第i 種群以及m 種不同資源中的第j 種資源,Monod 方程的微分形式如下:
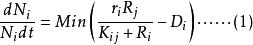 資源競爭
資源競爭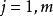 資源競爭
資源競爭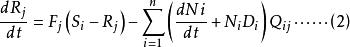 資源競爭
資源競爭式中, ri 為i 種群單位個體的最大生長率; kij 為資源j 限制下的種群i 的半飽和生長常數; Di 為種群i 的單位個體死亡率;Qij 為單位個體資源份額,即生產單位個體的種群i 所需的資源j 的量,其為產量的倒數; Rj 為資源j 的可得濃度; S i 代表為棲息地提供的資源j 的最大濃度,或是恆化培養器中資源j 的流入量; Fj 為資源j 的流通率或供給率; n 為種群數目;m 為限制性資源的數目。
公式(1) 表述的是一種群的生長率取決於環境中供給量最低的一種資源,即導致生長率最低的那種資源的濃度。公式(2) 表述的是資源j 的濃度取決於資源供給及資源消費。
Droop 競爭模型
Leham 等將Droop 方程及藻類對於資源吸收的米氏方程推廣至多種群間對於多種資源的競爭。對於n 個種群中的第i 種群及m 種不同資源中的第j 種資源,Droop 及米氏方程的微分形式如下:
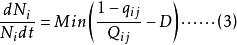 資源競爭
資源競爭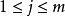 資源競爭
資源競爭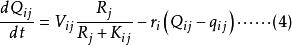 資源競爭
資源競爭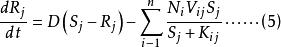 資源競爭
資源競爭式中, ri 為種群i 的最大生長率;qij 為種群i 貯存的資源j 的最小份額;D 為穩態下種群i 的死亡率; V ij為種群i 對資源j 的最大吸收速率;Kij 為種群i 吸收利用資源j 的半飽和常數;Qij 為種群i 對資源j 的內貯存份額; Ni 為種群i 的數量; S j 為資源j 的供給量;Rj 為資源j 的可得濃度;N 為種群數目; m 為限制性資源的數目。
公式(3) 表述的是種群的生長率取決於與相應的最小資源份額( qij ) 比值最小的那種資源的濃度( Qij) 。
R*-法則
當多個種群受一種資源限制時,令(1) 式等於零,得到一種群在平衡狀態下的資源需求值。
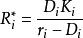 資源競爭
資源競爭式中,R* 為一種群在其生長率等於死亡率時的限制性資源的濃度。在多個種群競爭同一種非生物資源的情況下,如果它們的資源需求值作如下排列:R1*<R2*<R3*<R4*⋯⋯,那么根據Monod 方程,種群1 將能夠在競爭中淘汰其他種群,種群2 將能夠淘汰除種群1 以外的所有其他種群,依次類推。
這就是穩態下競爭單一非生物資源的R*法則。
資源比(resource-ratio) 假說
當多個種群受多種資源限制時,令(1) 式等於零,得到:
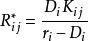 資源競爭
資源競爭式中,Rij*為種群i 的生長率等於死亡率時所需的資源j 的量。該式給出了在兩種資源限制下一個種群數量衰退、增長或維持不變的資源範圍。
實驗驗證
對資源競爭理論的實驗驗證和批評:
R* -法則及資源比假說已得到廣泛的實驗驗證。這些實驗的驗證對象從微生物、浮游植物到高等植物,及至浮遊動物。對資源競爭理論廣泛的實驗驗證是在實驗室內在微生物及浮游植物中展開的。R* -法則及資源比假說在這兩個生物類群中得到了很好的驗證。直接在自然條件下對R* -法則及資源比假說進行驗證的實驗是在高等植物間進行的,這方面的工作已廣泛開展並得到認可。資源競爭理論在對土壤中資源的競爭結果預測方面做得很好,但對於地上資源(如光) 的競爭結果預測似乎存在一些問題。在動物界,對資源競爭理論的驗證實驗僅見於浮遊動物,這些研究是在實驗室內在穩態條件下進行的,其結果支持R* -法則及資源比假說。
上述R* -法則及資源比假說中使用的種群在平衡狀態下的資源需求值R* ,皆得自Monod 方程。對於種群生長的Droop 模型,在資源供給及種群數量達到穩定的平衡狀態下,令(3) 式等於零,也將得到一個種群的資源需求值。
儘管資源競爭理論得到廣泛的實驗驗證,但這些實驗多數是在實驗室內高度可控的穩態條件下進行的,所以資源競爭理論是建立在眾多的假設前提條件之下的一種理論。這些假設的前提條件包括:系統的環境因子不隨時間、空間而改變;系統內不存在更高營養級的結構;種群內的生物個體之間無差異;生物體的生理特性不隨時間而改變;生物個體之間僅通過資源消費而發生相互作用;而且生物對不同資源之間的競爭消費不相關等。對一個自然生態系統來說,所有的這些假設可能都是錯誤的。依此理論,整個世界只會被少數幾個物種占據,但並非如此。事實上,改變任何一項上述所列假設條件都會使預測的共存和多樣性多起來。由此,許多學者嘗試打破系統的平衡性來驗證資源競爭理論,這一方面的工作主要是在浮游植物中展開的。
資源競爭理論的現狀及展望
儘管資源競爭理論已被許多的實驗證實,但這些實驗都是在單細胞生物間或多細胞生物間進行的。在自然界中,有些單細胞生物與多細胞生物也可利用並競爭同一資源,比如大型藻類與微藻,可以發生於同一水體,利用並競爭相同的資源。一般認為,單細胞生物的相對表面積(表面積∶體積) 遠大於多細胞生物,其對資源的吸收速度遠大於後者;而多細胞生物由於有器官或系統的分化,其對資源的貯存能力遠大於單細胞生物。由於這兩方面的差異,單細胞與多細胞生物間的競爭就變得更加複雜。關於單細胞生物與多細胞生物間的競爭是否符合資源競爭理論尚待進一步的實驗驗證,也許一個統一的既考慮環境中的資源又考慮體內資源貯存能力的模型正等待人們構建。
另外,對於資源競爭理論的進一步檢驗和發展,除了可以打破系統的平衡性(時間上的永恆性) 之外,許多理論學者還嘗試打破系統在空間上的均質性,或改變系統的營養級結構來進一步研究資源競爭的理論問題,從理論上提出了另外一些假說。例如,資源競爭理論的兩個假說都是以生物對非生物資源的消費和競爭為前提。事實上,在一個自然生態系統內,消費非生物資源的僅是初級生產者,食物鏈內的消費者皆以生物資源為消費對象。如果系統內的競爭者被天敵捕食,多種可能的競爭就會出現:這包括競爭取代、由捕食引起的競爭共存及優先性決定競爭等。為此有的學者提出了“防禦與競爭能力的交替換位( tradeoff s between defense and competitive ability) ”假說及“R*-和P*-法則”假說。假如系統的空間不均質,在空間結構上存在有孤立的島嶼或在空間分布上有一定規律但卻是分散存在的場所,針對這兩種不同的空間結構,Pacala等提出了“競爭2殖民交替換位( competitioncolonization tradeoff) ”假說和“拾遺者2機會主義者交替換位(gleaner-opportunist t radeoff ) ”假說; 除去系統的空間不均質外,系統內的光線也可能存在垂直分布,為此,Pacala 等提出了“光線梯度垂直分布(vertical partitioning of a light gradient) ”假說。
以上這些假說都未得到實驗檢驗,因此在嘗試驗證或修改這些假說方面還有大量研究工作要做。而且以上這些推測顯然還很不夠。比如,有一些學者嘗試打破在一個競爭種群內不存在年齡結構這一假設,對種群的年齡結構進行複雜的數學分析,建立了以高等植物為對象的競爭模型,只是這些模型僅局限於高等植物的生理特性。當然,期望在一個存在年齡結構的種群內得到可以通用的競爭法則很難,事實上,任何一項上述所列假設條件都很值得推敲。
更多的注意力應當放在資源競爭對生物進化的影響上。現有的研究給了人們一個令人吃驚的啟示,那就是雜合體在利用混合資源上的劣勢能夠推動生物資源利用分化的進程。當然,這個推論尚待進一步的理論和實驗研究。進化生物學家感興趣的許多生物生活史特徵都與生物吸收利用資源並將其傳遞給後代的能力有關。將資源競爭作為生物進化過程中一個明確的選擇壓力,不論在理論方面還是實驗方面,都有許多值得探討的東西。
