
原核生物介紹
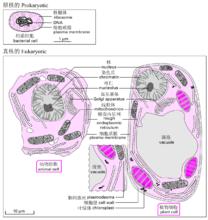 圖1 原核細胞與真核細胞結構的比較
圖1 原核細胞與真核細胞結構的比較與真核生物相比,原核生物顯得十分小巧(無論是細胞大小,還是其遺傳物質DNA分子),內部結構亦相對簡單——既見不到核,也見不到像線粒體、葉綠體這樣的細胞器(圖1)。不難理解,真核細胞必須用大的體積來裝填大的基因組,對複雜的基因組也需要進行更為有效有序的管控,這可能導致了細胞內部的模組化,譬如,將遺傳物質集中分布於用膜包裹的一個區域——形成所謂的核。在細胞核中,還有1個或多個圓球形的核仁,它是生產核糖體的機器。在現代生物學中,核被賦予了極為重要的地位,因為人們根據核的有無,將生物界區分為了兩大類——原核生物和真核生物。在植物細胞中,有一個稱之為葉綠體的細胞器專門用於光合作用。
共營模型
共營模型(syntrophic model)認為,古菌與細菌共生導致了含細胞核的真核細胞的誕生,但是,古菌與細菌均無細胞核(Hogan 2010) 。共營模型認為,與現代產甲烷古菌類似的某些古老的古菌,侵入並生活在類似於現代粘細菌的細菌體內,形成了早期的細胞核。古菌與真核生物在特定蛋白質(如組蛋白)基因的相似性被認為是支持以古菌為基礎的細胞核起源理論的證據。但筆者認為,共營模型並不能回答核是如何產生的問題。
其實,共營模型類似於解釋線粒體和葉綠體起源的內共生理論(endosymbiotic theory)。Konstantin Mereschkowsky於1905年提出葉綠體起源於內共生的猜想,而Ivan Wallin在1920s提出了關於線粒體起源的同樣的想法,之後葉綠體與線粒體內被發現含有自己DNA,雖然絕大多數基因都已丟失,譬如,核基因編碼了超過98%的線粒體蛋白質以及超過95%的葉綠體蛋白質,但要確定這些核基因有多少是來自線粒體或葉綠體的祖先可能並不容易。Margulis(1981) 使內共生理論得到普及,她認為,線粒體內共生起源於一種原核生物對另一種原核生物的攻擊,而不是所謂的吞飲(pinocytosis)或吞噬(phagocytosis)作用,因為這絕未在原核生物之間出現過;此外,內共生的螺旋體形成了真核生物的鞭毛和纖毛。此外,線粒體的原始宿主是厭氧的還是好氧的?因為含有複雜的氧化磷酸化系統,線粒體應該是在好氧環境下長期演化的產物。因此,如果它的宿主是厭氧的,那它至少通過與線粒體的共生獲得了對好氧環境更好的適應能力。
自演化模型
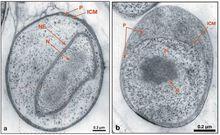 圖2 薄切片透射電鏡圖
圖2 薄切片透射電鏡圖自演化模型(autogenous model)認為原真核(proto-eukaryotic)細胞直接自細菌演化而來,並不需要通過內共生。證據來自一類專性好氧菌——浮黴菌(Planctomycete),它們具有清晰的胞內膜結構,其中,有一種稱之為 Gemmata obscuriglobus的出芽菌,其染色質被雙層的核膜所包裹(圖2a),類似於真核生物的核的結構,而斯氏小梨形菌( Pirellula staleyi)的核被單層的細胞質內膜ICM(圖2b)所包裹(Fuerst 2005)。但是,這一模型並未進一步解釋核實如何形成的。
圖2是 通過冷替代方式固定和準備的 Gemmata obscuriglobus(a)和斯氏小梨形菌( Pirellula staleyi)(b)的薄切片透射電鏡圖,NE—核膜,N—擬核,ICM—細胞質內膜,P—外室細胞質,P—pirellulosome(小梨形菌屬的一種膜細胞隔室)(引自Fuerst 2005) 。
起源模型
病毒性真核生物起源模型(viral eukaryogenesis model)認為,病毒感染原核生物導致了膜結合的細胞核與其他真核生物特徵的產生。證據是真核生物和病毒在大分子結構上存在一定相似性,譬如,線性DNA鏈、mRNA的加帽,以及與蛋白質的緊密結合(病毒的外套膜類似於組蛋白)。該假說的其中一種觀點認為,吞噬作用形成了早期的細胞“捕食者”,並隨之演化出細胞核(Bell 2001) 。另一種觀點則認為,真核生物起源於受到痘病毒感染的古菌,因為現代痘病毒與真核生物的DNA聚合酶具相似性(Villarreal and DeFilippis 2000 ,Takemura 2001 )。
外膜假說
外膜假說(exomembrane hypothesis)認為,細胞核是起源自演化出第二層外細胞膜的單個早期細胞,而包裹原來細胞的內膜則轉變成了核膜,並逐漸演化出精巧的核孔結構,以便於將內部(如核糖體亞基)合成的物質送出核外(de Roos 2006) 。
結構化假說
真核生物的DNA壓縮比
染色體結合有兩種蛋白質:組蛋白(一種低分子量的鹼性蛋白質)和酸性蛋白質。在真核細胞的有絲分裂過程中,與組蛋白耦聯的DNA分子的壓縮能力是十分驚人的(圖3)。DNA雙螺旋在每個組蛋白8聚體表面盤繞約1.75圈(長約140個鹼基對)構成核小體。相鄰核小體之間有長約50~60個鹼基對的DNA連線線,在相鄰的連線線之間結合了一個H1組蛋白分子。DNA似繩,組蛋白似珠,就像成串的珠子一樣,這就是染色體的一級結構,在這裡,DNA分子大約被壓縮了7倍。螺旋體是染色體的二級結構,每一周螺旋包括6個核小體,這裡的DNA被壓縮了6倍。螺旋體進一步螺旋化,形成超螺旋管,即染色體的三級結構,這裡的DNA被壓縮了40倍。超螺旋體再摺疊盤繞形成染色單體(四級結構),兩條染色單體組成一條染色體,這裡的DNA再被壓縮了5倍。這樣,DNA分子一共被壓縮了7×6×40×5=8400倍!
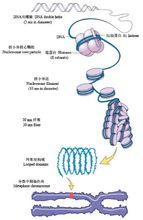 圖3 DNA壓縮成染色體的過程
圖3 DNA壓縮成染色體的過程原核生物與真核生物基因組大小的比較
再來看看這種DNA的壓縮與基因組演化之間的關係 。一般可用C值(C Value)來度量一種生物的基因組的大小,它指一種生物的單倍體基因組的DNA總量。細菌和古菌的C值(單位pg)的中位值約在10 –10 之間,而真核生物約在1-10之間,高約3個數量級。絕大多數C值落於中位數±1.5個數量級之間。此外,真核細胞略為原核細胞直徑的10倍。若考慮中位數及其上限偏移,以及細胞直徑一個數量級的差異,真核生物DNA的壓縮倍數大約應該是3.5個數量級(圖4),帶箭頭的紅色虛線表示C值中位數的演化軌跡,綠色虛線表示大多數真核生物C值的主要分布區間,問號表示該類生物起源的年代仍然存在爭議。這與現代真核生物的DNA壓縮比(packing ratio of DNA)驚人地一致!譬如,人的染色體中DNA分子伸展開來的長度平均約為幾個厘米,而染色體被壓縮到只有幾納米長(1 cm = 10 nm)。原核生物的C值與真核生物中的原生動物和真菌亦有交集,特別是與原生動物的交集更深。
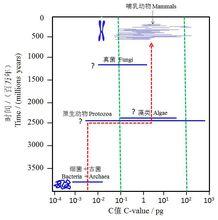 圖4 原核生物和真核生物基因組大小的比較
圖4 原核生物和真核生物基因組大小的比較壓縮與結構化假說
壓縮與結構化假說(packing and structurization hypothesis) 認為,細胞核源自基因組複雜化的誘導。在大氣逐漸氧化的背景下,地球上的生命(無論是形態結構,還是遺傳信息)加速了從簡單到複雜的演化歷程,可能主要由於DNA的複製錯誤或多倍化並在一定程度上疊加了不同種類之間的各種側向基因轉移方式以及內共生融合(endosymbioticfusion)等導致了一些原核細胞基因組的大型化。這一方面需要骨架蛋白的強化來支撐更大的細胞體積,同時通過個體生存的隨機性篩選,細胞內部逐漸結構化,從而形成了複雜的內膜系統——細胞器,被膜包裹的核及其在細胞分裂中的分離方式亦是這種結構化的產物。核的成型及有絲分裂的出現主要是為了滿足將巨大的DNA分子準確地分配到子代中去的需求,這裡,如何將長鏈DNA有效地壓縮(藉助組蛋白)成若干染色體以及如何將多個染色體同時分離(藉助紡錘體)是核演化的關鍵。核膜的形成雖然並非輕而易舉,但亦不會困難無比,膜有多種可能的來源,譬如,原核細胞分裂時DNA就得錨定在細胞膜上。其實,生命就起源自膜耦聯的光化學過程,細胞亦能產生各種各樣的膜。從本質上來看,包括核膜在內的細胞內膜系統就是為了實現對複雜生化系統進行秩序化管控,或者說,秩序化是通過細胞內部的模組化得以實現的。這雖然可視之為一種自演化模型,但在壓縮原理和結構化等的基礎上,詮釋了核演化的動因與本質(謝平2016)。
