
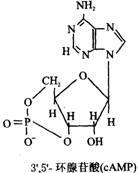
環狀核苷酸;核苷酸可在環化酶的催化下生成環式的一磷酸核苷。其中以3’,5’?C環狀腺苷酸(以cAMP)研究最多,它是由腺苷酸上磷酸與核糖3’,5’碳原子酯化而形成的,它的結構式如下圖所示。
 環狀末端核苷酸
環狀末端核苷酸環狀末端核苷酸
正常細胞中cAMP的濃度很低。在細胞膜上的腺苷酸環化酶和Mg2+存在下,可催化細胞中ATP分子脫去一個焦磷酸而環化成cAMP,使cAMP的濃度升高,但cAMP又可被細胞內特異性的磷酸二酯酶水解成5’?CAMP,故cAMP的濃度受這兩種酶活力的控制,使其維持一定的濃度。
現認為cAMP是生物體內的基本調節物質。它傳遞細胞外的信號,起著某些激素的“第二信使”作用。不少激素的作用是通過cAMP進行的,當激素與膜上受體結合後,活化了腺苷酸環化酶,使細胞內的cAMP含量增加。再通過cAMP去激活特異性的蛋白激酶,由激酶再進一步起作用。近年來發現3’、5’?C環鳥苷酸(cGMP)也有調節作用,但其作用與cAMP正好相拮抗。它們共同調節著細胞的生長和發育等過程。此外,在大腸桿菌中cAMP也參與DNA轉錄的調控作用。
2.核酸的化學結構(或一級結構)
核酸分子是由核苷酸單體通過3’,5’?C磷酸二酯鍵聚合而成的多核苷酸長鏈。核苷酸單體之間是通過脫水縮合而成為聚合物的,這點與蛋白質的肽鏈形成很相似。在脫水縮合過程中,一個核苷酸中的磷酸給出一個氫原子;另一個相鄰核苷酸中的戊糖給出一個羥基,產生一分子水,每個單體便以磷酸二酯鍵的形式連線起來。由許多個核苷酸縮合而形成多核苷酸鏈。如果用脾磷酸二酯酶來水解多核苷酸鏈,得到的是3’?C核苷酸,而用蛇毒磷酸二酯酶來水解得到的卻是5’?C核苷酸。這證明多核苷酸鏈是有方向的,一端叫3’?C未端,一端叫5’?C末端。所謂3’?C末端是指多核苷酸鏈的戊糖上具有3’?C磷酸基(或羥基)的末端,而具有5’?C磷酸基(或羥基)的末端則稱為5’末?C端。多核苷酸鏈兩端的核苷酸為末端核苷酸,末端磷酸基與核苷相連的鍵稱為磷酸單酯鍵。書寫多核苷酸鏈時,通常將5’端寫在左邊,3’端寫在右邊。但在書寫一條互補的雙鏈DNA時,由於二條鏈是反向平行的,因此每條鏈的末端必須註明5’或3’。通常寡核苷酸鏈可用右面的簡式表示(如右圖所示)。
述簡式還可簡化為pApCpGpUOH,若進一步簡化,還可將核苷酸鏈中的p省略,或在核苷酸之前加小點,則變為pACGUOH或pA?C?G?UOH。
3.核酸的性質
(1)一般性質
核酸和核苷酸既有磷酸基,又有鹼性基團,為兩性電解質,因磷酸的酸性強,通常表現為酸性。核酸可被酸、鹼或酶水解成為各種組分,其水解程度因水解條件而異。RNA在室溫條件下被稀鹼水解成核苷酸而DNA對鹼較穩定,常利用該性質測定RNA的鹼基組成或除去溶液中的RNA雜質。DNA為白色纖維狀固體,RNA為白色粉末;都微溶於水,不溶於一般有機溶劑。常用乙醇從溶液中沉澱核酸。
(2)核酸的紫外吸收性質
核酸中的嘌呤鹼和嘧啶鹼均具有共軛雙鍵,使鹼基、核苷、核苷酸和核酸在240~290nm的紫外波段有一個強烈的吸收峰,最大吸收值在260nm附近。不同的核苷酸有不同的吸收特性。由於蛋白質在這一光區僅有很弱的吸收,蛋白質的最大吸收值在280nm處,利用這一特性可以鑑別核酸純度及其製劑中的蛋白質雜質。
(3)核酸的變性和復性
①核酸的變性:是指核酸雙螺旋區的氫鍵斷裂,鹼基有規律的堆積被破壞,雙螺旋鬆散,發生從螺旋到單鍵線團的轉變,並分離成兩條纏繞的無定形的多核苷酸單鍵的過程。變性主要是由二級結構的改變引起的,因不涉及共價鍵的斷裂,故一級結構並不發生破壞。多核苷酸骨架上共價鍵(3’,5’?D磷酸二酯健)的斷裂稱為核酸的降解,降解引起核酸分子量降低。引起核酸變性的因素很多,如加熱引起熱變性,pH值過低(如pH11.5)的鹼變性,純水條件下引起的變性以及各種變性試劑,如甲醇、乙醇、尿素等都能使核酸變性。此外,DNA的變性還與其分子本身的穩定性有關,由於C?DC中有三對氫健而A-T對只有兩對氫鍵,故C+G百分含量高的DNA分子就較穩定,當DNA分子中A+T百分含量高時就容易變性。環狀DNA分子比線形DNA要穩定,因此線狀DNA較環狀DNA容易變性。
