介紹
染色體倒位(inversion)是由於同一條染色體上發生了兩次斷裂,產生的斷片顛倒1800後重新連線造成的。如果倒位發生在染色體的一條臂上,稱為臂內倒位(paracentric);如果倒位包含了著絲粒區,則稱為臂間倒位(pericentric)。臂內倒位不改變兩個臂的長度,要用染色體顯帶技術才能識別;臂間倒位則使兩個臂的長度出現增減,即使未作染色體顯帶處理也可觀察區分。
特點
 染色體倒位
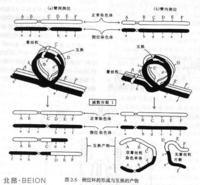
染色體倒位在減數分裂中,正常的染色體同倒位染色體之間發生交叉互換,會使配子染色體上某一區段缺失或重複,從而造成染色體異常,導致子代出現異常性狀。這是因為倒位雜合子在減數分裂時,兩條同源染色體不能以直線形式配對,一定要形成一個圓圈才能完成同源部分的配對,這個圓圈稱為倒位環(inversion loop)。
一個倒位雜合體如果著絲粒在倒位環的外面,則在減數分裂後期會出現“斷片和橋”的現象,即一條染色單體的兩端都有一個著絲粒,成為跨越兩端的“橋”,同時伴隨一個沒有著絲粒的斷片。“橋”在染色體移向兩極進入子細胞時被拉斷,造成很大缺失;斷片則不能進入子細胞的核內,所以由此形成的配子往往是死亡的。
一個倒位雜合體如果著絲粒在倒位環的裡面,在環內發生交換後,雖然不會出現“橋”和斷片,但也會使交換後的染色單體帶有缺失或重複,形成不平衡的配子。這種配子一般也沒有生活力
倒位的套用
由於倒位可抑制重組,人們就利用此特點將它套用於突變檢測和致死品系的建立。
1.ClB技術
早在二十年代Muller就利用果蠅建立了一系列檢測基因隱性突變和致死突變的技術,ClB就是其中諸多方法之一,雖然現在已有一些先進的分子遺傳學的手段但ClB技術仍不失為一種較為簡便的方法,更值得借鑑的是Muller將基礎理論套用於實踐的一些思路,其構思精巧和靈活令人嘆服。
ClB技術中的C表示交換(crossover),Muller選擇了X染色體上存在倒位的果蠅,目的是抑制交換,這樣分析時就可以排除重組的因素。l是指致死基因(lethat),即在倒位的X染色體上已帶有一個隱性致死基因;B是指棒眼(Bar)基因,為顯性突變。ClB方法如下(圖22-14):將ClB品系的雌果蠅與待測的雄果蠅(例如事先經X光照射)交配,在F1代中雌果蠅中應有兩種表型:棒眼及非棒眼;在雄果蠅中應只有一種野生型可以存活,而另一種帶有ClBX染色體的雄果蠅不能存活,因Y染色體上沒有L基因的正常等位基因,使l表達而致死。再選擇帶有棒眼的F1雌果蠅和野生型雄果蠅雜交,產生的F2代也有兩種類型,一種棒眼(ClB/+),另一種非棒眼(?/+)。棒眼雌果蠅一定帶有一條ClB染色體,可以用來用下一輪檢測。非棒眼的雌果蠅雖帶有和待測的X染色體,但由於另一條X染色體是野生型的,所有的基因都正常,所以即使待測的X染色體上發生了隱性突變或致死隱性突變都不有顯現出來。在雄果蠅中帶有ClBX染色體的應不能存活,餘下的一類雄果蠅是帶有一條待測的X染色體和一第Y染色體,若經X-射線照射產生了隱性突變即可直接在這一類雄果蠅中直接顯出來,若產生隱性致死,那么所有的雄果蠅都不能成活。
選擇雄果雄作為待測的對象是因為(1)一隻雄果蠅可以和多隻雌蠅交配,這樣篩選突變(F1代)的樣本就增加了。(2)雄果蠅是完全連鎖的,不會發生交換。雌果蠅用ClB有以下的優點:(1)用C(倒位)可抑制交換,使得結果明確,不受重組的干擾,(2)B(棒)眼作為一種標記易區分兩類雌蠅,加將ClB品系分離出來;(3)l使一半無待測X染色體的雄蠅致死,這樣觀察結果較為方便。
2.保存帶有致死基因的品系——平衡致死品系(balancedlethalsystem)
致死基因的存在是可以供各種研究和檢測(如ClB技術)用的,但獲得一個致死突變並非易事,由於往往是隱性致死,即使是產生了也還得費很大的氣力去檢測和分離,因此人們總想建立一個致死品系,這樣用起來就較為方便。首先提出平衡致品系的構想是Muller。1918年他在實驗室里發現了一種顯性突變叫做蠣殼翅(beaded,Bd)(翅的前緣有扇形皺褶)Muller將Bd突變的果蠅相互雜交,結果後代有兩種類型,蠣殼翅和野生型,但比例不是3:1而是2:1,表明Bd基因可能純合致死,使Bd/Bd這種純合類型致死。但有一次突然出現了意外的情況,蠣殼翅果蠅互交的後代都是蠣殼翅。Muller對於偶然出現的現象並不放過,他對此做了一個假設,即在Bd等位基因的附近產生了一種新的致死突變,而且染色體存在著倒位,抑制了可能發生的重組。人們就利用此原理建立了一系列的平衡致死品系,這樣它們互交的後代都是平衡致的,即兩個不同的致死基因反式排列在一對同源染色體上,且有倒位的存在抑制了重組,無須選擇就能保持真實的遺傳,使致死品系得以保存。
