孢子綱簡介
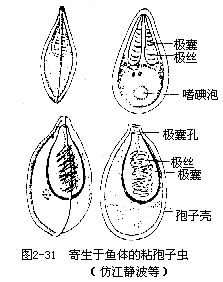
原生動物門的一綱。營寄生生活,沒有運動細胞器,但在生活史中一定階段,具有偽足或鞭毛。有兩個寄主或一個寄主,都產生孢子(spore),藉以傳播,習稱孢子蟲。具有兩個寄主的種類,其整個生活史都在寄主體內進行,孢子在寄主體內發育成熟,這樣的孢子無殼;只有一個寄主的種類,孢子必須在寄主體外發育,這樣的孢子具有較厚的外殼,這是對外界不良環境產生的一種適應性。約有3500種,分布廣,人體、家畜、家禽及其他動物等都有孢子蟲寄生,危害很大。寄生在人或脊椎動物的紅細胞內的孢子蟲,如寄生人體的瘧原蟲,寄生家畜的焦蟲等,一般稱血孢子蟲,都有兩個寄主,一為人或脊椎動物,一為節肢動物,如蚊、蟬等。有的孢子蟲主要寄生在魚類的體表、鰓、皮下及內臟各組織中,影響魚類的發育生長。它們產生的孢子結構複雜,孢子內一般具有1~4個小囊,囊內有一團細絲,稱為極囊和極絲;孢子具外殼,脫離寄主後,被新寄主吞食或遇到新寄主時,極囊中的極絲即放射出,刺在寄主體上,逐漸形成小結節,即在其內發育,形成許多裂殖子,成熟後再產生配子,這類孢子蟲稱為粘孢子蟲。有的寄生在桑蠶或蜜蜂腸壁細胞中,稱蠶微粒子和蜂微粒子,可使蠶僵化,使蜜蜂瀉痢。它們的孢子很小。一般無極囊,只有一條極絲,少數種類有一個極囊。這類孢子蟲稱微孢子蟲。此外還有寄生在草食性獸類的橫紋肌內的肉孢子蟲;寄生在兔的膽管或腸上皮中的球蟲。細菌也是靠孢子繁殖的。代表動物
——間日瘧原蟲(Plasmodium vivax Grassi&Feletti)
瘧原蟲能引起瘧疾,這種病發作時一般多發冷發熱,而且是在一定間隔時間內發作,有些地方叫“打擺子”或“發瘧子”,是我國五大寄生蟲病之一。
已描述的瘧原蟲有50多種,其中寄生在人體的瘧原蟲主要有4種:間日瘧原蟲(P.vivax)、三日瘧原蟲(P.malaria)、惡性瘧原蟲(P.falciparum)和卵形瘧原蟲(P.ovale)。瘧原蟲的分布極廣,遍及全世界。在我國以間日瘧和惡性瘧為最常見;卵形瘧在我國極少發生。在東北、華北、西北等地區主要為間日瘧,三日瘧較少。惡性瘧主要發生在我國西南,如雲南、貴州、四川、海南島一帶。過去所說的瘴氣,其實就是惡性瘧。
這4種瘧原蟲的生活史基本相同。現以間日瘧為例說明瘧原蟲的形態和生活史。
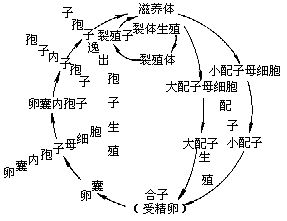
間日瘧原蟲有2個寄主:人和按蚊。生活史複雜,有世代交替現象。無性世代在人體內,有性世代在某些雌按蚊體內,借某些按蚊傳播。
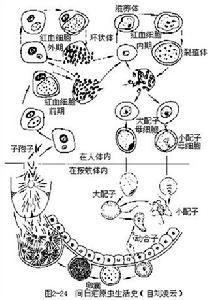
(一)在人體內(進行裂體生殖)瘧原蟲在肝細胞和紅血細胞內發育。有些學者把在肝細胞發育分為紅血細胞前期和紅血細胞外期。有些學者把紅血細胞前期稱為紅血細胞外期。在紅血細胞內發育包括紅血細胞內期及有性時期的開始(配子體形成)(圖2—24)。
紅血細胞前期(pre-erythrocytic stage);當被感染的雌按蚊叮人時,其唾液中瘧原蟲的長梭形子孢子(sporozoite),隨唾液進入人體,隨著血流先到肝臟,侵入肝細胞內,以胞口攝取肝細胞質為營養(這時稱滋養體trophozoite),逐漸增大,成熟後通過復分裂進行裂體生殖(schizogony)。即核首先分裂成很多個,稱為裂殖體(schizont),裂殖體也以胞口攝取肝細胞質為營養,然後細胞質隨著核而分裂,包在每個核的外邊,形成很多小個體,稱裂殖子或潛隱體(merozoite或crypto-zoite)。當裂殖子成熟後,破壞肝細胞而出,才能侵入紅血細胞。因此把瘧原蟲侵入紅血細胞以前,在肝細胞里發育的時期稱為紅血細胞前期(即病理上的潛伏期)。在此期中一般抗瘧藥對瘧原蟲沒有什麼作用。此期在間日瘧一般為8~9天,惡性瘧原蟲需6~7天。這是屬於短潛伏期的,但也有長潛伏期的。我國學者江靜波教授等,近些年來對河南省間日瘧原蟲多核亞種(P.vivax multinucleatum)長潛伏期的研究,經三位參加該項工作的同志獻身作試驗,用人工感染的蚊蟲叮咬,結果證明該間日瘧原蟲在人體內分別為312、321和323天。這是我國首次以實驗的方法證明長潛伏期間日瘧在我國的存在,並獲得了長潛伏期具體時間的精確記錄。
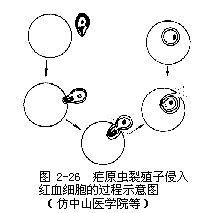
裂殖子成熟後,脹破肝細胞,散發在體液和血液中,一部分裂殖子可被吞噬細胞吞噬,一部分侵入紅血細胞,開始紅血細胞內期的發育。一些學者認為,還有一部分又繼續侵入其他肝細胞,進行紅血細胞外期發育。
紅血細胞外期(exo-erythrocytic stage):由於此時在紅血細胞內已有瘧原蟲,因此相應地稱為紅血細胞外期。用抗瘧藥物治療,紅血細胞內瘧原蟲雖被消滅,但外期的瘧原蟲並沒有被消滅,當它們在肝細胞內行裂體生殖所產生的裂殖子出來以後,侵入紅血細胞可使瘧疾復發,因此外期的存在是瘧疾復發的根源。最近許多學者認為紅血細胞外期尚未完全證實,而認為瘧疾愈後的復發,是由於子孢子進入人體侵入肝細胞後,一部分立即進行發育(也稱為早髮型或速髮型子孢子tachysporozoite),引起初期發病。其餘的子孢子處於休眠狀態(也稱為遲髮型子孢子bra-dysporozoite),經過一個休眠期,到一定時候休眠的子孢子才開始發育,經裂體生殖形成裂殖子,侵入紅血細胞後引起瘧疾復發。
這些裂殖子經過幾次裂體生殖周期以後,或機體內環境對瘧原蟲不利時,有一些裂殖子進入紅血細胞後,不再發育成裂殖體,而發育成大、小配子母細胞。在間日瘧,大配子母細胞(雌)較大,有時較正常紅血細胞可大一倍,核偏在蟲體的一邊,較緻密,瘧色素也較粗大。小配子母細胞(雄)較小,核在蟲體的中部,較疏鬆,瘧色素較細小(惡性瘧的配子母細胞形狀如臘腸,雌的兩端稍尖,核較緻密,雄的兩端鈍圓,核較疏鬆)。這些配子母細胞在人體內如不被按蚊吸去,不能繼續發育。在血液中可能生存30~60天。
瘧原蟲對人的危害很大,它能大量地破壞紅血細胞,造成貧血,使肝脾腫大,近年來發現間日瘧原蟲也能損害腦組織,嚴重地影響人們的健康甚至造成死亡。解放前,危害極為嚴重,如雲南南部一
孢子綱的主要特徵
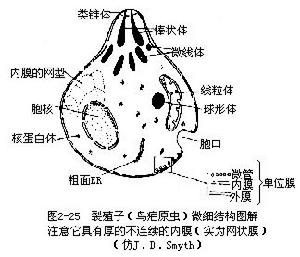
孢子綱的動物都是營寄生生活的,無運動器,或只在生活史的一定階段以鞭毛或偽足為運動器,這可說明孢子蟲與鞭毛蟲和肉足蟲的親緣關係。孢子綱動物很多具有頂複合器(apical com-plex)結構,因此近年來有學者將具有頂複合器的孢子蟲另列為頂復體門(Apicomplexa)(原孢子綱的微孢子蟲、粘孢子蟲也分別立為門)。頂複合器包括類錐體、極環、棒狀體、微線體等結構。對這些胞器的功能還不很了解,有人認為類錐體、棒狀體和微線體等與寄生蟲侵入寄主細胞有關。營養方式為異養。生活史複雜,有無性世代與有性世代的兩個世代的交替,這2個世代多數在2個寄主體內進行,無性世代在脊椎動物(或人)的體內,有性世代在無脊椎動物體內進行。也有些種類在同一寄主體內進行。無性生殖是裂體生殖。有性生殖是配子生殖,其後為無性的孢子生殖。也有人將配子生殖與孢子生殖合稱為有性生殖。其生活史大多經過以下幾個時期,圖解見《普通動物學第四版》50頁最上方圖。
孢子綱的重要類群
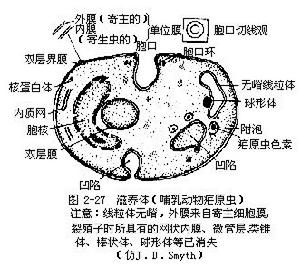
球蟲
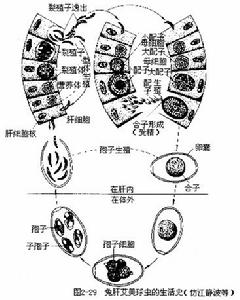
:這類孢子蟲多寄生於脊椎動物消化器官的細胞內。生活史與瘧原蟲的基本相同,不同的是,它只寄生在一個寄主體內,卵囊必須在寄主體外進行發育。孢子有厚壁。主要寄生於羊、兔、雞、魚等動物體內。如兔球蟲,寄生在肝膽管上皮細胞的為兔肝艾美球蟲(Eimeria stiedae)(圖2-29),寄生在兔腸上皮細胞的有穿孔艾美球蟲(E.perforans)等。據調查我國至少有9種兔球蟲,這幾種球蟲一般多混合感染,對家兔危害很大,尤其對斷奶前後的幼兔更為嚴重,有時可引起家兔大量死亡,對養兔業是很大威脅。其生活史與瘧原蟲基本相同。即兔誤食了卵囊(感染階段)後,子孢子在小腸內從囊內出來,侵入肝膽管的上皮細胞或腸上皮細胞內發育成滋養體,進行裂體生殖。過一段時期後產生大小配子母細胞,進行配子生殖,形成合子,在其外分泌厚殼,稱為卵囊,卵囊隨糞便排出體外。在合適的外界條件下卵囊發育,核分裂形成4個孢子母細胞,每個孢子母細胞外分泌外殼,成為4個孢子,每個孢子內又分裂成為2個子孢子,即每個卵囊內有8個子孢子。在此階段的卵囊,如被另一兔吃下就可被感染,或者重複感染。卵囊對外界條件的抵抗力很強,根據實驗,用濃度低於1%的石炭酸、高錳酸鉀、來蘇兒以及飽和鹽水和鹼水等不能殺死卵囊,但用80℃以上水處理可使卵囊迅速死亡,因此用開水洗刷兔籠及用具或用80℃~100℃高溫處理是預防此病的有效措施。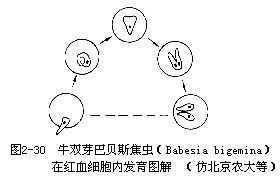
血孢子蟲
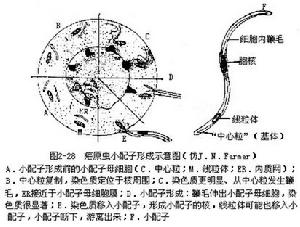
:在其生活史中經過兩個寄主。裂體生殖時期寄生在脊椎動物或人體內(血液中或細胞中),配子生殖和孢子生殖是在吸血的節肢動物(蚊或蜱)體內。由於其整個生活史在寄主體內進行,所以孢子無殼,如瘧原蟲。又如在我國巴貝斯焦蟲(Babesia)和泰勒焦蟲(Theileria)①70000034_0059_0對家畜均有危害,可引起家畜患焦蟲病。焦蟲有許多種,通常寄生在家畜的紅血細胞內,蟲體呈圓形、環形、梨形(單個或成對)等不同形態(圖2—30)。不同的家畜各有其一定的焦蟲致病,彼此互不感染,其中以牛焦蟲種類最多,其它家畜的較少。病原體通過硬蜱傳播。當被感染的硬蜱吸取家畜血液時,焦蟲即可進入家畜的紅血細胞(泰勒焦蟲先進入淋巴細胞和組織細胞中繁殖,再進入紅血細胞內寄生),然後以二分裂或成對出芽法進行繁殖(圖2—30)。當紅血細胞破裂後,蟲體再侵入其它的紅血細胞。如此反覆進行,破壞大量紅血細胞。嚴重時一部分血紅蛋白由腎排出,成為血尿。焦蟲在蜱體內發育繁殖較複雜,有些具體過程尚不完全清楚或無一致結論。有些種類在蜱內發育一段時間後,一部分蟲體侵入蜱的卵巢,隨著蜱卵的形成,被包在卵內,並藉此傳遞給下一代,當蜱的幼蟲到家畜體上吸血時又傳給家畜。或者不經過卵傳遞,蜱在幼蟲或若蟲階段吸血感染焦蟲,焦蟲即在蜱體內發育繁殖,等蜱長成若蟲或成蟲吸血時再傳給家畜。焦蟲對家畜危害較大,有的死亡率可高達90%。