卵軸
正文
生物體以及卵和細胞都具有物質分布不等或生理特性不同的兩端,這種性質稱為極性。根據卵內物質(如卵黃、色素顆粒、核糖體等)沿卵軸分布的差異,可把卵分為動物性半球和植物性半球。如兩棲類成熟卵的細胞質較集中的區域稱為動物性半球,卵黃較集中的區域稱為植物性半球。動物性半球和植物性半球的中央分別稱為動物性極和植物性極。卵的極性一般可從其外形(如昆蟲和頭足類的卵)及細胞核的位置、色素的分布(如棘皮動物和兩棲類)、卵的某種特殊附屬結構(如硬骨魚的卵膜孔)的位置等清楚地分辨出來。也有一些動物卵的極性不易分辨。極性是卵子在卵巢內的發生過程中形成的。各類動物卵巢結構不同,影響卵子極性形成的因素也可能不同。有人認為極性的形成同生長中的卵母細胞的兩端在卵巢內的處境(如血液供應、質膜通過微胞飲作用攝取卵黃磷蛋白前體,鄰接的營養細胞或濾泡細胞的分布和功能活動等)不均衡有關。也有人認為同卵母細胞細胞核的極化和中心粒的位置有關。在減數分裂的偶線期,染色體簇集在核一端,朝向中心粒,最初的卵軸就是通過核──中心粒的軸。對於極性形成迄今還沒有一種統一的解釋。脊椎動物的卵多數是沿卵軸呈輻射對稱的。成體兩側對稱的體型是在胚胎髮育過程中逐漸建立起來的。蛙卵受精後,與精子穿入點相對的動物性半球區域出現色素較淡的灰色新月可作為兩側對稱的最初的外部標誌。第一次卵裂時,沿卵軸通過灰色新月中央的分裂面,相當於未來成體的左右對稱面。原腸形成開始時,胚孔的背唇最先在原來灰色新月所在的位置上出現,因而確定了胚胎的背腹方位。灰色新月的位置與精子穿入點有一定的關係,定位受精實驗證明:大約90%的卵上,灰色新月在離精子穿入點相對的135°~180°的扇形區內出現。因此,精子穿入點基本上可作為預測未來的背唇和胚胎背腹軸位置的根據。也有人認為卵在受精前已存在某種不穩定的兩側對稱結構,精子穿入的作用是使這種結構穩定下來。
80年代初對爪蟾受精卵的研究表明,精子入卵後引起的卵質流動和胚泡破裂後所含物質在背方的重分布,可能在卵的背腹軸和兩側對稱的建立中起著重要的作用。
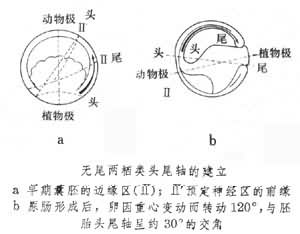
隨著原腸形成的完成和中軸器官的形成,胚胎出現了頭尾軸。胚胎的頭尾軸與卵軸往往是不一致的,例如,在蛙類由於原腸形成及形態發生運動,尤其是內胚層捲入內部和神經板的形成,使胚胎重心改變而發生轉動。此時頭尾軸大約是水平的,以致動物性極向前腹方移動約120°,處於頭尾軸的腹方,所以原來的卵軸和胚胎頭尾軸呈現約30°的交角 (見圖)。卵軸和胚軸有一定的關係,其交叉角度隨物種不同而異。
 卵軸
卵軸