
由來
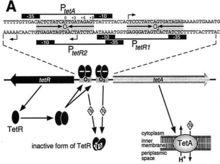
Ramos等將原核生物調控因子分為16個家族:LysR、AraC/XylS、TetR、LuxR、LacI、ArsR、IcIR、MerR、AsnC、MarR、NtrC (EBP)、OmpR、DeoR、Cold shock、GntR和Crp,其中參與抗生素生物合成的調控因子廣泛存在於TetR家族。TetR家族是對TetR蛋白的遺傳和生化全序列特徵的基礎上加以命名的。TetR蛋白控制tet基因的表達,這些產物能夠抑制四環素的產生。TetR家族成員在DNA結合域方面有高度的序列相似性。TetR是四環素溢出泵基因的敏感調控子,其作為TetA的決定子,TetR蛋白依據其胺基酸序列的相似性分為八類(TetR A到E,G,H和J),它們有超過40%的同源性。儘管RA1質粒編碼的有218個胺基酸的TetR(D)的結構已經闡明,且和TetR(B)有63%的同源性,但是到目前為止具最好特徵的TetR蛋白是Tn10編碼的有207個胺基酸的TetR(B)。
結構特點
 tetR
tetR大多數細菌的轉錄調控子含有兩個結構域:一個是接受信號的結構域,一個是DNA結合的結構域。結構分析表明在原核轉錄子中大多數的DNA結合模組通常是螺旋-轉角-螺旋(HTH)的結構。在原核生物中約有95%的轉錄因子採用HTH模組結合它們的靶DNA序列。
TetR二聚體是由兩個完全相同的單體組成,由轉角和環連線的10個a螺旋摺疊而成。TetR單體的三維結構主要通過疏水的反向螺旋維持穩定。TetR二聚體的總體結構可以分為:位於每個單體N末端的兩個DNA結合區域;由二聚化和配體結合的調控核心區。DNA結合區由螺旋a1,a2,a3及對稱螺旋a1',a2',a3'構成,螺旋a4和a4'再將其連線到由螺旋a5,a10和對稱螺旋a5',a10'組成的調控核心區。調節區負責二聚化和當二價陽離子存在的情況下每個單體的結合位點。螺旋a5,a8,a10和對應螺旋構成核心區支架,並且它們的結構在整個TetR構象中是最為保守的。
