農桿菌介導法起初只被用於雙子葉植物中,近年來,農桿菌介導轉化在一些單子葉植物(尤其是水稻)中也得到了廣泛套用。
一 農桿菌介導的基因轉化方法
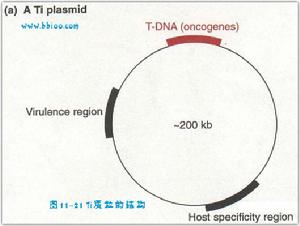
(一)農桿菌的Ti質粒與T-DNA的整合機制幾乎所有雙子葉植物都容易受到土壤農桿菌感染,而產生根瘤。它是一種革蘭氏陰性土壤桿菌(A. tumefaciens)。其致瘤特性是由Ti(tumor-inducing)質粒介導的。農桿根瘤菌之所以會感染植物根部是因為植物根部損傷部位分泌出酚類物質乙醯丁香酮和羥基乙醯丁香酮,這些酚類物質可以誘導Vir(Virulence region)基因的啟動表達,Vir基因的產物將Ti質粒上的一段T-DNA單鏈切下,而位於根瘤染色體上的操縱子基因產物則與單鏈T-DNA結合,形成複合物,轉化植物根部細胞。T-DNA上有三套基因,其中兩套基因分別控制合成植物生長素與分裂素,促使植物創傷組織無限制地生長與分裂,形成冠癭瘤。第三套基因合成冠癭鹼,冠癭鹼有四種類型:章魚鹼(octopine)、胭脂鹼(nopaline)、農桿鹼(agropine)、琥珀鹼(succinamopine),使農桿菌生長必需的物質。
1. Ti質粒的結構
在發現根瘤農桿菌誘發冠癭瘤的本質是Ti質粒後,Ti質粒便成為冠癭瘤形成基因鑑定與分析的主要研究對象。
Ti質粒大約在160~240kB之間。其中T-DNA大約在15kb-30kb。Vir基因區在36kb左右。除此之外,Ti質粒上還存在Con區(region encoding conjugation)和Ori區(origin of replication)。
T-DNA上共有三套基因和左右兩個邊界,LB和RB是長為25bp的末端反覆重複順序,在切除及整合過程具有重要意義。
tms由兩個基因組成:tms1(iaaM)和tms2(iaaH)
tmr由一個基因組成iptz:
tmt由若干基因構成,合成稀有胺基酸衍生物,稱為opines。它有三個成員:
octopine=精氨酸與丙酮酸的縮合物
Napaline=精氨酸與-酮戊二酸的縮合物
Agropine=谷氨酸與二環糖的縮合物
據此可將Ti質粒分為三大類,感染的植物誘導合成這些有機鹼,但不能利用它們,其分解酶基因在Ti質粒上,分解產物為胺基酸和糖類,供根癌農桿菌使用作為氮源及碳源。
2.T-DNA的整合機制:
T-DNA的詳細整合機制尚不清楚,但有幾個環節是明確的:
T-DNA切除由Vir區編碼的特異性內切酶完成,分別在LB和RB的第三個鹼基和第四個鹼基之間產生缺口,並形成單鏈T-DNA。
T-DNA的LB和RB在整合中的作用是不對稱的,RB順序與整合有關,而LB無關。
T-DNA的整合可以是單拷貝的,也可以是多拷貝的,成串聯形式排列,但整合位點的特異性尚未確定。
(二)Ti質粒轉化植物細胞的戰略
1 . Ti質粒的改造
有以下理由使天然的Ti質粒不能作為表達載體使用:
a. 生長在培養基上的植物轉化細胞產生大量的生長素和分裂素阻止了細胞再生長為整株植物,因此,必須除去生長素和分裂素基因。
b. 有機鹼的合成與T-DNA的轉化無關,而且可能會影響植物細胞生長,因為有機鹼合成大量消耗精氨酸和谷氨酸,因此必須去除有機鹼合成基因(tmt)
c. Ti質粒約為200kb,重組操作非常苦難,也很難找到單一的酶切位點。
d. Ti質粒不能在大腸桿菌中複製,為了使重組質粒DNA的大量擴增,須添加入大腸桿菌複製子。 加入植物細胞的篩選標記,如neor基因,使用植物細胞啟動子及末端polyA化信號,加入多聚人工接頭以利於外源基因的克隆。
植物中一般不存在質粒,為利用農桿菌的Ti質粒,發展了共整合系統和雙元載體系統,避免了在大的Ti質粒上進行分子重組操作的困難。
2. 植物細胞轉化的共整合系統
T-DNA克隆在大腸桿菌質粒上,含有E.coli的選擇標記和植物選擇標記Kmr。首先在E.coli中篩選重組分子,然後將重組質粒轉化到農桿菌中,質粒與Ti質粒上的同源序列發生同源重組,將外源基因整合到Ti質粒上,用於侵染植物細胞。T-DNA重組分子整合到植物細胞染色體DNA上,Kmr篩選轉化細胞。
3 . 植物細胞轉化的雙元系統
目前T-DNA轉化植物細胞的標準方法是雙元系統,即穿梭質粒。插入外源基因的重組穿梭質粒直接轉化含有Ti質粒的根瘤農桿菌,經篩選後直接感染植物細胞。與共整合系統所不同的是,含外源基因的質粒可在農桿菌內自主複製並保留下來。農桿菌侵染植物細胞後,植物的創傷信號啟動Ti質粒上的Vir基因,隨後將穿梭質粒的T-DNA切割下來,轉移到植物細胞中。
(三)改良植物性狀的策略
基因克隆技術提供了一種新的改良植物的方法,它可以直接的改變植物的基因型。有兩種策略可以套用。
1) 基因附加:通過添加1個或多個基因改變植物的性狀。
2) 基因扣除:利用基因工程技術使一個或多個植物已經存在的基因失活。
滅活植物基因是通過反義技術來實現的。將外源基因反向的連線到載體中,當基因轉錄成mRNA後,與正常的mRNA是反向互補的。我們稱這種反向互補為反義RNA,縮寫為asRNA。
反義RNA阻止原有基因表達的機理尚不明了,但可以肯定,正義和反義RNA之間的雜交與這一事件有關,可能雙鏈的RNA很快被核酸酶降解 ,或者反義RNA阻止了核糖體與正義RNA的結合。
