耐輻射奇球菌簡介
耐輻射奇球菌( Deinococcus radiodurans, DR)是地球上已知物種中最耐電離輻射的生物之一。它是1956 年由美國科學家Anderson 等首先從輻照滅菌後仍然發生變質的肉類罐頭中分離出來的。該細菌屬於極端微生物,對電離輻射、紫外線、乾燥、強氧化劑和一些化學誘變劑等各種DNA 損傷介質的致死和突變效應顯示驚人的抗性,被譽為“地球上最頑強的細菌”。
耐輻射奇球菌的形態和生長特性
 耐輻射奇球菌
耐輻射奇球菌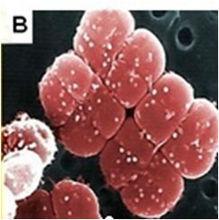 耐輻射奇球菌
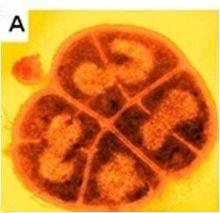
耐輻射奇球菌 耐輻射奇球菌形態圖
耐輻射奇球菌形態圖耐輻射奇球菌的菌落呈圓形,直徑約在1~2 μm之間,好氧,能產生粉紅色色素,不產孢子。單克隆的菌苔呈凸狀,表面光滑,在指數生長期,約90%的細菌呈二聯體存在;隨著細胞分裂,生長後期形成四疊體;在穩定期,絕大多數細菌呈四疊體。其最適生長溫度是30℃,在37℃時生長速度最快。當溫度低於4℃或高於45℃時,細胞停止生長。
耐輻射奇球菌的細胞結構
耐輻射奇球菌有一個不尋常的細胞壁結構,雖然它的細胞壁成分和一般的蘭
氏陰性細菌相似,但是由於它的肽聚糖層比較厚,難以脫色,因此實際革蘭氏染
 耐輻射球菌細胞膜模式圖
耐輻射球菌細胞膜模式圖色呈陽性。其細胞壁可以劃分為一個14-到20-nm 的肽聚糖層和一個未知的分層結構,在電鏡下,至少可以分為六層,最裡面一層是細胞膜,緊挨著細胞膜是含有肽聚糖的細胞壁,細胞壁上有很多孔,也稱為多孔層,壁上的這些孔對細胞有重要的生理學意義。第三層是可以分成無數的細小的區域(分區層),第四層是外膜,第五層是電子緻密區;第六層由規則排列的六角形的蛋白亞單位組成(S 層, 或六角排列的中間體層),也有一些耐輻射奇球菌外面還有一層厚的多糖外膜。但是在細胞分裂時,只有細胞質膜和肽聚糖層參與形成細胞間隔斷,其他的各層僅作為外鞘,在分裂的子細胞分離時,包繞在子細胞外面。
細胞外膜和細胞質膜在脂類組成上一樣,但沒有發現其他菌中普遍存在的脂
多糖。耐輻射球細胞壁的脂肪酸組成也是與眾不同,它既不含有羥基脂肪酸、類
脂A 以及庚糖,也不含有多不飽和脂肪酸、環丙基和分支脂肪酸,而含有一個
由15-, 16-, 17-, 以及18-碳飽和或單不飽和脂肪酸組成的混合物。和其他細菌另一個明顯的不同是缺乏磷脂,耐輻射球細胞膜的脂類中,43%是含有烷基胺的磷酸甘油酯 。
耐輻射奇球菌的基因組結構
耐輻射奇球菌為多基因組,穩定生長期細胞中至少含有4 個拷貝的基因組,而在分裂旺盛的指數生長期則可達10 個以上拷貝。冗餘的基因組拷貝數給細胞提供了一個遺傳信息儲蓄庫,使其通過同源重組修復DNA 雙鏈斷裂更有優勢。Levin-Zaidman等描述了在耐輻射奇球菌類核中的一個不同尋常的環狀結構。高度濃縮的類核結構對輻射抗性有貢獻,因為這種結構使得耐輻射奇球菌在產生DSB 後也能維持DNA 線狀連續性。基因組構象與DNA修復的關係顯然很重要,然而Zimmerman 等人和本實驗室的研究結果則認為類核環形結構對耐輻射奇球菌的電離輻射抗性並不是必須的。
 耐輻射奇球菌 耐輻射奇球菌基因組構成圖
耐輻射奇球菌 耐輻射奇球菌基因組構成圖1999 年,White 等在Science 上公布了耐輻射奇球菌野生型菌株R1 的全基因組序列。整個基因組由4 個環狀分子組成,包括兩條染色體(染色體I 為2.65 Mb、染色體II 為412 kb) 、一個巨型質粒(177 kb) 和一個小質粒(45. 7 kb),總共3284156bp,平均G+C 含量66.6%。整個基因組編碼3187 個開放閱讀框(Open reading frames,ORFs),平均大小約937 bp,覆蓋了整個基因組的91%。其中約1/3的基因不能從現有資料庫中找到同源物(1002 個)或是功能未知的(511 個)和複製有關的基因主要位於染色體I 和II。
耐輻射奇球菌的DNA 損傷抗性
耐輻射奇球菌是最具輻射抗性的生物體之一。指數生長期的耐輻射奇球菌對γ-射線表現出極強的抗性,存活的最高劑量是15 kGy(若在生長後期,則它們的存活率更高些,可達到17 kGy),是人體細胞耐受力的3000倍,且沒有任何產生突變的證據。在8kGy 的電離輻射下,耐輻射奇球菌仍保持10%的存活率,而大腸桿菌在0.15 kGy照射劑量下才能達到10 %的存活率。另外,指數生長期的耐輻射奇球菌在在5kGy的電離輻射下,對數生長期的耐輻射奇球菌細胞不受任何影響;其D37(存活率為37 %時的輻射劑量)為6.5 kGy左右,而在該電離輻射劑量下大腸桿菌(E. coli)和枯草桿菌(Bacillus spp.)都不能生長。
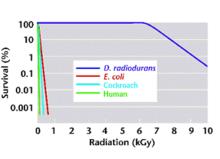 輻照條件下不同生物的生存狀況
輻照條件下不同生物的生存狀況在紫外線抗性上,耐輻射奇球菌可以在高達1000J/m的UV處理後存活下來,在500J/m2的劑量下,它的生存沒有影響,而大腸桿菌的存活則急劇下降。對數生長期的耐輻射奇球菌的UV抗性大約是大腸桿菌的33倍左右。
耐輻射奇球菌具有很強的抗乾燥能力,在一個乾燥器記憶體放六年後,仍能保
10%的存活率。科學家研究證明了耐輻射奇球菌的輻射抗性和乾燥抗性的關係。他們將野生型和41株對輻射敏感的耐輻射奇球菌同時放在乾燥器內,六周后檢測他們的存活率,結果表明對輻射敏感的菌株對乾燥同樣敏感。表明耐輻射奇球菌抗乾燥機制和抗輻射機制有關。
耐輻射奇球菌經大劑量γ-射線照射(15 kGy)後染色體基因組產生約150~200 個雙鏈斷裂(Double-strand break, DSB)的DNA 片段,約3000 個單鏈片段(SSBs)和至少1000 個損傷的鹼基位點。其中DSB 是DNA 損傷中最致死的形式。儘管所有生物都具有DNA 修復機制,但在多數物種中只有很少一部分DSB 能被修復。DNA 脈衝場凝膠電泳實驗表明,電離輻照後耐輻射奇球菌染色體基因組由原來的2.65×10 kb 泳帶變為50 kb 或更小的一條帶,這些產物對一般細胞都是致命的,而受損傷的耐輻射奇球菌基因組卻能在幾十小時之內完全修復。
耐輻射奇球菌極端抗性機制
耐輻射奇球菌對輻射、乾燥、化學試劑毒性等極端環境有明顯的耐受性,歸因於其在細胞結構,遺傳信息,代謝調節,損傷修復等方面形成了一套完善的抗性機制。
▪ 耐輻射奇球菌的自我保護
在結構上,耐輻射奇球菌細胞擁有多方面的保護,如外面有一層較厚的細胞壁,細胞膜內含有大量的類胡蘿蔔素。另外,基因組內編碼多種清除活性氧的蛋白,包括三種過氧化氫酶,至少三種超氧化物歧化酶和過氧化物酶,有利於防範因電離輻射,UV 、強氧化劑和乾燥等造成的活性氧損傷細胞。如電離輻射的間接作用能在細胞內產生大量的自由基,超氧化物歧化酶(SOD)、過氧化氫酶(CAT)和過氧化物酶(POD)能有效清除細胞內超氧陰電離自由基(Oˉ)、逕自由基(·OH)和過氧化氫(HO)等對細胞毒害作用極強的活性氧自由基。生物體內的SOD、CAT 和POD 3 種酶活性的高低和所含的其他保護物質的量,在一定程度上也決定著對輻射抗性的強弱。在正常條件下,耐輻射奇球菌細胞中SOD 的活性比對照大腸桿菌高出6 倍,用Mn處理穩定期的細胞,其SOD 的活性又比未處理的高出3倍。耐輻射奇球菌細胞中CAT 的活性更是比對照的大腸桿菌高出30 多倍。耐輻射奇球菌的SOD、CAT 和POD 等保護酶的高含量成了其輻射抗性極強的一個原因。
耐輻射奇球菌菌體內大量合成以deinoxanthin為主要產物的類胡蘿蔔素類胡蘿蔔素。Deinoxanthin具有特殊結構,該色素由一個六元環和一條長不飽和碳鏈組成,環上含有一個羥基(2-OH)和酮基(4=O),長鏈末端尾部還具有一個羥基(1′-OH),其結構與現在已知的主要色素種類如番茄紅素(lycopene)和β-胡蘿蔔素(β-carotene)等均存在明顯差異,自由基清除能力比同類色素均要強很多,因此認為該特殊類胡蘿蔔素作為高效的抗氧化劑,參與了菌體內抗氧化體系,對該菌的極端抗性機制具有重要貢獻。
▪ 冗餘的遺傳信息
耐輻射奇球菌細胞內有多個基因組拷貝,一般在穩定期的細胞內有4個拷貝的染色體,而活化的細胞內則有4-10條染色體。多拷貝的基因組提供了冗餘的遺傳信息,有利於減少因輻射造成的遺傳信息丟失,從而有利於DNA修復,使其在抗逆性中扮演重要角色。
▪ 染色體間重組
經過5kGy劑量輻照處理後,耐輻射奇球菌的每個染色體上可產生150個左右的DNA雙鏈斷裂。細胞內的核酸外切酶可以作用於DNA斷裂處,迅速破壞DNA末端,造成序列信息的丟失。儘管如此,耐輻射奇球菌還是有能力將這些碎片重新組合成完整的染色體。斷裂是隨機產生的,由此在每條染色體上造成的遺傳信息缺失也是隨機的,因此儘管每一條染色體都有斷裂,但是不同染色體間斷裂的分布是不同的,如果細胞可以介導染色體間碎片重排的話,理論上,碎片是足夠修復形成一個完整的染色體。實驗證明在耐輻射奇球菌的四條染色體間,輻照可以誘導產生600多個交聯,其中大約有三分之一是單向的,說明耐輻射奇球菌可能是通過將染色體片段連線到一起來重建其完整染色體。
▪DNA 修復的有序調節
在真核細胞中,DNA損傷可以抑制細胞的DNA複製。在原核細胞中,只有當DNA損傷阻礙了沿著模板移動的DNA聚合酶時, DNA複製才會停止。然而在耐輻射奇球菌中,也存在著輻射誘導的DNA複製抑制現象。實驗證明在耐輻射奇球菌染色體中,損傷引起的複製暫停時間和劑量相關,並且停頓的時間總是超過修復引起停頓的DNA損傷時間,表明耐輻射奇球菌的DNA複製對細胞內的DNA損傷是很敏感的,細胞內有一套檢測DNA損傷修復程度的機制發揮作用,並可給複製系統傳導修復的信息。也就是說耐輻射奇球菌內有一套類似真核生物的可調節的檢測點,可以控制DNA複製以及後面的細胞分裂。研究人員從遺傳學上進一步證明了耐輻射奇球菌DNA損傷檢測點的存在,他們鑑定了耐輻射奇球菌突變體SLR2、SLR4和SLR5,這些突變體的輻射抗性沒有改變,倍增時間也和野生一樣長,不影響生長,但是輻照後恢復明顯變慢,形成菌落需要更長的時間。
▪DNA 降解
電離輻照後,耐輻射奇球菌中迅速發生廣泛的染色體DNA 降解。這種降解可能是由細胞內的DNA 單鏈或雙鏈斷裂引起的。對大多數物種來說,伴隨著DNA 降解造成的遺傳信息丟失是輻射誘導死亡的一個重要原因。然而,研究表明耐輻射奇球菌的DNA 降解是其修復的一部分,降解的過程和輻照劑量相關,劑量越大,降解持續的時間越長,染色體DNA 的損失也就越多。DNA 降解速率與劑量無關,每分鐘大約有0.1%的染色體被降解。耐輻射奇球菌限制DNA 降解的程度是細胞對電離輻射致死效應的一種保護。
▪ 損傷 DNA 的外排
染色體DNA降解伴隨著損傷DNA片段由細胞內向細胞外的排出。DNA損傷早期,在耐輻射奇球菌的細胞質及培養液中均發現DNA損傷片段,初期降解產物是寡核苷酸,長約2 kb鹼基對,隨後降解為核苷,可快速地從細胞中排出。這一階段被認為是耐輻射奇球菌DNA損傷修復的第一階段,稱“細胞清除期”。游離核苷酸輸出的程度與輻照的劑量呈正相關。儘管向外釋放DNA損傷和損傷抗性的關係還沒有詳細的研究,但是向外排除損傷是修復鹼基損傷的一種有效的機制,也代表一種存活策略,可能有兩方面的作用:其一、向外排除損傷核苷酸可以減少突變水平,阻止損傷的鹼基在隨後的合成中再次利用;其二、向外排除鹼基是協調DNA修覆信號的一個環節。耐輻射奇球菌中有多種UvrA同源蛋白,其中的某些UvrA蛋白除能識別DNA損傷外,也許還參與了損傷核苷酸的排出。
▪ 錳含量和蛋白質氧化
Daly等研究了高濃度Mn(Ⅱ)的作用,發現Mn的聚集可以增強 D.radiodurans的抗輻射能力。當 D. radiodurans處於缺乏Mn的培養條件下時,它們的抗離子輻射能力也就下降。不論是否有Mn的存在,該菌在特定輻射劑量下形成的DNA雙鏈斷裂數是相同的,因此Mn並不能阻止DNA損傷。但Mn存在時,該菌種更能忍受高輻射劑量導致的細胞損傷。
大多數輻射產生的DNA損傷是活性氧的產生導致的。細胞內的Mn通過清除活性氧來實現保護作用。例如,乳桿菌( Lactobacillus plantarum)缺少過氧化物岐化酶的保護酶,取而代之的是細胞內20-25mM濃度的Mn離子。由於 D. radiodurans的雙鏈斷裂似乎並不受Mn的影響,因而活性氧的清除一定是保護大分子而不是DNA,猜想Mn離子的富集阻止了過氧化物並和輻射蛋白損傷活性氧產生有關。如果存在這種情況,那么沒有聚集足夠Mn離子的細菌可能死於離子輻射引起先於DNA損傷的蛋白損傷。已通過實驗證實,Mn的確可以保護蛋白免受自由基的攻擊。經輻照處理後,基於小分子Mn(2+)的抗氧化保護機制保持了DNA修復和複製蛋白高效性。
除此之外,Mn離子濃度的增加可以提高 D. radiodurans基因組的濃度。當在生物體DNA溶液環境中加入多價陽離子時,DNA就會被濃縮,這是因為陽離子中和了骨架中磷酸基團相斥的部分。
▪ 特有的 DNA 保護和修復因子
經歷電離輻射的細胞內反應是非常複雜的。電離輻射可誘導產生多種類型的DNA 損傷,包括雙鏈斷裂(DSB)、單雙鏈斷裂(SSB)、DNA-蛋白交聯和不同類型的鹼基損壞等。細菌的DNA 修復包括一些部分冗餘的途徑,被認為是一種適應。耐輻射奇球菌擁有全套的細菌DNA 修復系統如鹼基切除修復、核苷酸切除修復、鹼基錯配修復和同源重組修復等。許多科學家認為耐輻射奇球菌極強的抗輻射能力與其本身具有完善而高效的DNA修復系統有關。但在耐輻射奇球菌中是否存在SOS 反應和非同源末端重組(NHEJ),一直是研究人員所疑惑的。至今尚未有確鑿的證據來證明這兩條途徑對耐輻射奇球菌的抗性有作用。
利用基因組學和蛋白組學,篩選到了一批輻照和乾燥處理後被大量誘導表達基因,其中一部分為功能未知的特有基因,如 ddrA、 ddrB、 ddrC、 ddrD和 pprA等。現在探明一些重要未知基因的功能。DdrA蛋白具有末端保護的功能,結合在DNA單鏈的3’端的,防止它們被核酸酶的降解。通過對斷裂DNA末端的保護,細胞能夠保護基因組DNA 直到環境適合細胞的生長和DNA 的修復。 ddrA突變株在營養豐富的基質中培養的細胞輻射敏感性只有略微的提高。而如果將細胞輻射後使其處於飢餓狀態,那在五天之後其生存能力只為野生型的1/100。DdrB蛋白有具有和DdrA蛋白類似卻不同的保護機制。而PprA蛋白被認為能在體外促進DNA雙鏈斷裂的連線。這些蛋白可能對該細菌的超強抗性均具有重要貢獻。
在耐輻射奇球菌中新發現了一個能夠促進DNA損傷修復的多效開關因子PprI。該蛋白突變株對電離輻射、紫外線輻射和絲裂黴素C等DNA損傷介質均極其敏感。可以在DNA損傷後激活 recA、 pprA與其他DNA修復基因的轉錄與翻譯,並可以大幅度地提高過氧化氫酶的活性,啟動了包括了脅迫回響途徑、轉錄翻譯途徑、能量和物質代謝途徑、抗氧化途徑等在內的DNA損傷回響和細胞生存網路,是耐輻射奇球菌抗輻射必需的。因此,在該菌中存在著別具一格的DNA損傷回響機制,使得其具有非凡的NA修復能力和耐輻射抗性 。
