外形特徵
新鮮蟲體呈肉紅色肥厚泡狀體,兩側較深,中間無色透明,伸縮活動,體形變化不一。固定後,體橢圓形。體表滿布體棘,群生型,呈尖刀狀並列成簇。棘群含棘數目在兩盤問最常見為10支,在兩睪間最常見為20支。但在不同地區、不同宿主,略有差異。由豬肺取出的蟲體棘簇叢狀,排列整齊,亦不易脫落。消化系統:口呈漏斗狀,在口吸盤內通入咽中,後接食管,食管常因蟲體伸縮大小不定
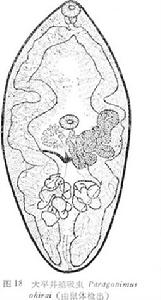
卵巢:從同一來源的囊蚴感染不同宿主,所獲蟲體的卵巢形態有所不同:在貓體者,卵巢中心體與腹吸盤稍有距離,其前端內側分支有的和腹吸盤外側重疊。中心體有為棒狀,前端有6個分支,分支遠端膨大,表面有小的隆凸;在豬、鼠體者,成蟲的卵巢中心體粗大,遠端有較多而細的分支;有的中心體為塊狀,前端伸出大的隆凸,其上又有小隆凸如生薑狀;有的卵巢中心體為棒狀,前端分出較多的分支,分支前端又有第二或第三分支,形成珊瑚狀,後一種形態在寄生鼠類的蟲體中尤為多見。
子宮:位於卵巢的對側,其基部達腹吸盤與睪丸間,當子宮內蟲卵豐盈時,子宮管變粗短如結腸迂迴重疊,形成髻狀,其前緣伸達腹吸盤水平或超越腹吸盤前緣水平,後端與同側睪丸前部重疊,外側與腸管重疊或越過腸管之外,內側遠端在卵黃管前越過蟲體縱中線到達對側。當子宮內蟲卵排空時,子宮管即變為細長。生殖孔在腹吸盤後有一定距離。
卵黃腺的發育、分布和子宮內蟲卵的充盈程度相一致。當子宮內蟲卵豐盈時,除蟲體中間排泄囊的一條狹窄空隙區無色透明和生殖器官外,大部被卵黃腺充占,濾泡緻密,卵黃管也相應出現,左右兩橫卵黃管在蟲體正中匯合形成圓形或棱形的卵黃囊,呈棕黃色。勞氏管位於生殖孔與子宮之間。
兩睪丸傾斜並列於蟲體後部1/3的腸管第三彎曲內。每睪丸由基部伸出4--6分支,一般常為5支。不同宿主的蟲體,其睪丸的形態也有差異:睪丸分支呈大小球狀,排列集中,面積較小,寄生於貓體的蟲體,多屬此類型;睪丸為塊狀,一側有粗短隆凸,寄生於豬、犬體的蟲體多為此類型;睪丸分支為裙帶狀,末端膨大,分支向多方面伸出,面積較大,寄生於鼠類的蟲體,多為此類型。輸出管由睪丸向前伸出,左右管匯合形成輸精管,後者粗短,在腹吸盤後方通入貯精囊,貯精囊與受精囊形狀、大小相近似。受精囊位於貯精囊內側和輸卵管、梅氏腺的後方,平均大小為0.161×0.102。
蟲卵棕黃色,卵圓形,兩側對稱,卵殼厚薄均勻,卵蓋明顯,後端不增厚。從兩隻大白鼠怖內的蟲卵分別為84.84×51.32與81.20×48.56微米,從貓肺內取出的蟲卵大小為84.84×47.65微米,從豬肺內取出的蟲卵則為70.41×47.86微米。
生活習性
終末宿主 國內自然感染宿主有貓、犬、豬、溝鼠等。在日本方面有溝鼠、豬、鼬鼠、狸、獾等。貓可作為理想的實驗動物。第一中間宿主在上海為兩種擬沼螺(Assiminea violacea 與 A .colombeliana)及釘螺(Oncomelania hupensis)在廣州的兩種擬沼螺(A.lutea 與A.violacea)和浙江的放逸短溝蜷(Semisulcospira libertina)(即川卷螺)亦人工感染成功。第二中間宿主在上海自然感染的有兩種相手蟹(蟛蜞)(Sesarma(H.)dehaani及S.sinensis)和天津厚蟹(Helicetridens tientsinensis);在遼寧營口地區為一種相手蟹(s.(H.)dehaani)和天津厚蟹(H.tridens tientsinensis)。另外,中華絨鉗蟹(Eriocheir sinensis)及日本蝲蛄(Cambaroides japo-nicus)人工感染成功。
生活史(圖19) 作為中間宿主的螺和蟛蜞孳生在每晝夜潮汐淹沒6—8小時的江河沿岸,潮汐水位高時在水底活動,退潮後即在濕潤地面活動,幼螺和幼相手蟹的活動範圍,退潮後距離水流較近淹沒的時間較長,隨著日齡的增長與水流距離較遠,在地面活動的時間則較長,到夏秋季節則多在近岸活動。長大的相手蟹,在水淹沒時多向岸邊活動,稻田窪地,庭院下水道,房舍常可見到。
蟲卵:在28—30℃時,18天發育成熟,毛蚴孵出。毛蚴壽命在夏季為24小時左右,溫度低則壽命稍長。當毛蚴與螺宿主相遇則蜂擁而上,從螺體露出的軟體部分鑽入螺體,毛蚴與螺接觸10分鐘約80%的毛蚴消失不見,15分鐘後,活動力強的毛蚴已近完全消失。毛蚴進入螺體變為胞蚴,在夏季毛蚴感染擬沼螺(A.violacea)後,自24小時至30日間皆可自螺體檢出胞蚴。早期胞蚴呈圓形或橢圓形,蚴體漸長即變長,伸縮形成節狀,6日齡時體長為0.135—0.173,體節寬為0.039和0.67。第10日齡的胞蚴體長0.177x 0.072,其體內已出現早期的母雷蚴7—8個。15日齡的胞蚴體大小為0.339—0.517×0.073—0.172,蚴體出現棕色縱顆粒條紋兩條,此條紋可隨胞蚴伸縮活動分向兩側展開或合併。20日後,戰形的母雷蚴在螺體內出現。當螺與毛蚴數目比例1:2集中感染時,陽性螺體內含有的胞蚴數,最少為1個,最多為18個。30日齡後的胞蚴體內母雷蚴變少,棕色顆粒增多,體色變深,活動能力低弱,出現衰老現象,並逐漸消失。
母雷蚴:擬沼螺(A.violacea)感染16—20日後,母雷蚴即在螺體頭部和中部的消化道周圍淋巴中出現。一個陽性螺含有母雷蚴的數量,一般在50個以內。雷蚴體表光滑無色透明,咽和原腸清晰,後端常向一側彎曲。因發育程度不同大小不一。幼小的母雷蚴後端平齊向內凹陷,7個72日齡活體染色未加壓標本平均大小為0.367×0.0127,頭部0.042,咽0.032,原腸0.089×0.066,體內含有子雷蚴8—10個左右。感染後32曰,螺體內有大量雷蚴,一般在50個以上,常見為母、子雷蚴同時出現。子雷蚴一般較母雷蚴為大,內部結構更清晰明顯,原腸短小,多呈棕黃色,蚴齡一年以上者固定染色標本平均大小為0.627×0.105,頭部0.049,咽0.042,原腸0.078×0.068,體內生殖細胞15個左右(11—20個)。從釘螺體內剖出的子雷蚴固定染色標本大小為1.037×0.136,頭部0.0339×0.0446,咽0.042×0.033,原腸0.127×0_339,體內有細胞團4個左右和成熟的尾蚴。從川卷螺體內剖出的子雷蚴,蚴體較大,固定染色標本大小為1.547×0.425,頭部0.029×0.096,咽0.088×0.088,原腸0.118×0.072,體內含有發育成熟的尾蚴21--44個和細胞團分布於蚴體前端到後端。值得注意的是大平並殖在川卷螺體內發育的子雷蚴體大,含有尾蚴和細胞團數量多,尾蚴形態呈正常伸展狀態。而在擬沼螺體內發育,據剖檢的感染兩年以上的陽性螺體內只有子雷蚴66--77個和大量不同發育程度的未成熟尾蚴,此結果表明了大平並殖的胞蚴和母雷蚴到一定階段即衰老消失。尾蚴是由存留的子雷蚴繁殖產生,直到螺死亡。
尾蚴:毛蚴侵入螺體內後,開始排尾蚴的時間,排蚴數量及規律,尾蚴壽命皆和季節氣溫有非常密切關係。陽性螺於春初五月中旬前後開始陸續排出尾蚴,炎夏6、7、8月份排蚴數達到全年最高峰,入冬11月份前後排蚴停止。根據一個毛蚴感染一隻A.violacea和20個以上的毛蚴感染的A.violacea的觀察結果表明,其排尾蚴規律比較,除停放一周以上後的第一次排蚴在數量上有增高傾向外,一般未出現顯著差異。陽性螺每次逸出的尾蚴最少為1個,最多為1000個以上。尾蚴為微尾型,自螺體逸出時,體形細長,群.集於粘液塊中扭動,約10分鐘左右,粘液液化,尾蚴在水底自由蠕動。尾蚴壽命在春秋季節一般為24--48小時,和溫度成反比,近死亡的尾蚴伸脹為橢圓形。從A.colombeliana逸出的尾蚴用5%福馬林溶液固定標本測量大小為:體部0.157X 0.081,尾部0.017x0.019,口吸盤0.029×0.031,口吸盤背面椎刺0.013×0.005,腹吸盤0.021×0.021,排泄囊呈三角形或呈葫蘆狀,生殖原基塊狀,位於體後部,蚴體後部和尾部具有小毛,尾內有細胞5、6個。
以逸出尾蚴人工感染相手蟹,在解剖鏡下看到尾蚴在水底向蟹體方向移動,當接觸蟹體時即附著其上,尋覓柔軟部位,以頭部錐刺尖端固定,後端不停鏇動,短時內即進入蟹體。在上海地區,夏秋季節,尾蚴與幼小相手蟹接觸一小時,3.5小時,16小時與48小時後解剖檢查結果,尾蚴絕大多數是從相手蟹的螯肢,腿關節尤其第三腿的第三關節鑽入。蟹類宿主的脫殼期和斷肢再生新肢,是蟹類宿主解除對尾蚴入侵的預防機能階段,如果在其附近有陽性螺存在或從螺體逸出尾蚴粘液團沖至其附近,則可造成感染。尾蚴鑽入相手蟹後,部分死亡,部分尾蚴尾部脫落。大多數尾蚴仍帶尾部向肝臟移行,感染後第13天即到達肝臟,個別尾蚴停留於鰓、胸肌、螯肢、腿、殼下等處,發育形成囊蚴。感染後20日齡的囊蚴,形態結構基本形成。20個囊蚴的平均大小為0.287×0.244,囊壁厚度為3.2微米。20個脫囊後尾蚴的平均大小為0.326×0.196,口吸盤為0.057X 0.061,腹吸盤為0.055×0.064,咽為0.031×0.0316,椎刺為0.010X 0.002,腸管有三個彎曲,體披小棘,此時被大白鼠吞食後可檢獲蟲體。 囊蚴20日齡後即不再見增大,唯囊內後尾蚴結構更為清晰鮮明。囊壁增厚,後尾蚴縮伏囊內。34日齡囊蚴有的體內已出現紅色顆粒。65日齡20個囊蚴的平均大小為0.286×0.232,囊壁厚度約為5--6微米,脫囊後尾蚴體披小棘,20個平均大小為0.357×0.235,口吸盤直徑0.052,椎刺為0.001×0.003,腹吸盤0.064×0.074,咽0.026,腸管有三個大彎曲,排泄囊從腸管分叉後緣伸達體後端。
動物實驗表明,家貓在吞食大平並殖囊蚴後,第40日或42日即發育成熟,糞便中開始出現蟲卵,蟲體在貓體生活的時間較長,在解剖自然感染和10個人工感染10個月以上的貓時,從未發現過死亡的蟲體、變性蟲囊,或有卵無蟲等現象,且檢獲的蟲體較大、姿態自然、體色新鮮。從自然感染的豬和人工感染豬及犬肺內解剖所見蟲體和蟲囊與寄生貓肺者相近似,應當認為貓、犬、豬是此蟲的正常終末宿主。
與人的關係到目前為止,國內尚未見有在人體發現成蟲和確診有本蟲感染的報導。在上海,曾對在黃浦江水系沿岸本蟲流行區居住的有吃相手蟹史的成人和學齡兒童進行廣泛的痰液和糞便檢查,未發現有並殖吸蟲蟲卵。豬受感染後,影響其生長發育,嚴重時可使養豬事業造成損失。
分布範圍
中國分布:遼寧上海、台灣、廣州。國外分布:日本、
物種研究
1939年宮崎在日本的兩種相手蟹(蟛蜞)(Sesarma intermedia 及S.dehaani)的肝臟檢出並殖吸蟲囊蚴,人工感染大白鼠獲得成蟲,經鑑定為新種,命名為大平並殖吸蟲。 國內陳心陶在1933年研究怡樂村並殖的同時,亦在廣東的相手蟹(Sesarma[H.]dehaani)中發現此種囊蚴,當時為解決這種具厚壁的囊蚴及它們所變成的成蟲是否即怡樂村並殖,或另為一個獨立種,曾進行比較研究,亦認為它們是屬於不同種的(陳心陶,1960)。宮崎(1943)在我國的吳淞發現蟹類(s.[H.]dehanni)被大平並殖囊蚴感染,解放後安耕九(1954)進一步發現其第一中間宿主為A.violacea。1948年安耕九等又在我國上海北郊家貓肺部檢出大平並殖成蟲。1950年在同地豬肺內又檢獲此蟲。並對當地兩種相手蟹(s.dehaani及S.sinensis)進行檢查,在肝臟均發現有並殖囊蚴,人工感染大白鼠,獲得大平並殖吸蟲成蟲。此外,發現貓、豬、溝鼠、晏鼠和狗有自然感染。
1966年李得垣等在遼寧營口地區亦發現此蟲。
