
概述
達爾文的進化論已經創立140餘年了,在其誕生之初是作為一種假說被提出來的,除達爾文本人從對一些植物,動物形態的觀察得出的推論外,這些年學術界完善了鈾鉛測年法對化石證據的排序和基因證據。達爾文在《物種起源》書中論及化石時,標題為“不完美的地質記錄”。他承認在當時的化石研究中並未有證據顯示有物種間過渡類型的存在,並指出這可能是最易於檢驗而又具有殺傷力的反進化論的理由。他看到了進化論的理論缺陷,並希望後人能予以驗證、完善。時至今日,在當今任何一本生物學雜誌上,已經找不到任何質疑進化論的論文了。SCOTT和COLE在八十年代初,檢索了當時的4000多種學術刊物,未發現任何一篇反進化論的論文,在68種與生物起源有關的學術期刊中,也未發現任何一篇是質疑進化論的。GEORGE W. GILCHRIST在1997年調查了世界上最大的五種期刊數據索引,也未發現反進化論或非進化論的論文。
發展
SCOTT和COLE的工作還發現,在1985年提交的135000篇論文中,確有18篇論文是反進化論的和非進化論的,這都是質疑進化論理論缺陷的。這更加給人一種印象:進化論並非確立於自身學說的科學性和完美程度,而是確立於眾多分子生物學的數學模型。進化論並非KARL POPPER意義上的“經驗科學”(EMPIRICAL SCIENCE),而是一個數學模型,純數學的證據的雜合體。
事實上,我們今天科學研究所發現的東西,已經足以讓人們重新最佳化進化論的不足了。一些人在思維定式的驅使下,自覺或不自覺地成為盛行理論的衛道士,而喪失了獨立思考的能力,這是不符合理性的科學精神的。一個真正的科學家,應正視舊理論的缺陷及其面臨的挑戰,並勇於擺脫束縛。只有這樣,科學才能向前發展,人類才能向前推動。
觀點
一. 理論簡介
(一)比較解剖學
進化論者通過動物的器官在形態和功能方面的類比,定了所謂的同源器官,並由此說明在進化樹中某一譜系的動物該器官在進化中發生的形態與功能的變化是自然選擇的結果。首先,同源器官的定義就非常牽強,你必須首先承認動物是進化的,才能找到同源器官。因此這絕不能算做進化論中的一個證據,而只能是一個推論。即我們只能說因為進化論正確,所以進化樹中某一譜系中的動物存在同源器官,而不能說同源器官的存在證明進化的存在。現代基因學和遺傳學誕生後,對生物體形態與功能的關係在更本質的層次--基因及分子水平有了嶄新的認識。形態和功能只是表相,它們是由基因決定的,相同的形態可能對映於完全不同的基因。如果認為從鳥類的翅到哺乳動物的前肢是進化的話,那么它們的基因也應表現為對映於形態相同程度的進化。但實際上並非如此,如果現在仍有人試圖從表面現象說明問題,只能被認為是膚淺的。
(二)古生物學
1. 凌亂的化石證據
無論是進化論者還是反進化論者,都希望古生物化石能輔證自己的觀點 。如果說達爾文時代缺少化石記錄的話,那么到今天為止,人類已收集了數以萬噸記的化石,是否已證明進化論的正確了呢?事實上依然不能。CHICAGO的FIELD MUSEUM OF NATRURAL HISTORY的古生物學家RAUP本是進化論者,但後來不得不放棄自然選擇學說,而支持幸運者生存說。他說該館擁有現有已知化石物種的近五分之一,可是已經確證的中間過渡類型的化石比達爾文時代還少。現有化石記錄混亂不堪,人為按進化論組成的譜系漏洞百出,根本不能說明問題。進化論者們所使用的分類方法及標準又不同,因此對某一化石的斷代也經常爭論不休,得不到一個統一的結論。
進化論者常用的一個較為完備的進化譜系證據是脊椎動物序(VERTEBRATE SEQUENCE),包括從魚類到兩棲類,爬行類,鳥類,哺乳動物,直至人的進化。從魚類到兩棲類,曾發現一種叫RHIPIDISTIAN的魚化石,其骨骼特徵類似於兩棲類,因此被認為是魚類到兩棲類的過渡類型。但當另一種叫COELACANTH的活化石發現後,這種觀點被立即推翻,因為這兩種動物在進化譜系中的親緣關係是極近的,但研究表明,COELACANTH並無任何上岸的可能性。曾經轟動一時的始祖鳥(ARCHAEOPTERYX),曾被視為進化論的鐵證,因為它即具有爬行動物的特徵,又具有鳥類的特徵而被視為一個著名的過渡類型。後來發現,始祖鳥所具有的爬行類特徵,如爪和牙齒,在現代鳥類中也存在,如南美洲OPISTHOCOMNSH-OATZIN 和非洲的TOURACOCORYTHAIX。也就是說這個證據並不能用於判定鳥類與爬行類的過渡類型。另外,JENSEN早在1977年《SCIENCE》上發表的論文指出,與始祖鳥同一地層出土的化石被懷疑為近似於現代鳥類動物的股骨,因此,始祖鳥即非“始祖”,只是種間類型。
在從猿到人的進化過程中,拉瑪古猿(RAMAPITHECUS)具有重要意義。它的下頜骨兼具人類與猩猩的特徵,牙床結構類似於人,不具有猩猩的門齒與犬齒,上下頜的距離與頜骨的長度又近似於猩猩,這被視為是進化論的一個鐵證。然而後來在非洲發現的一種狒狒(THEROPITHECUSGLADE)與拉瑪古猿具有相同的牙齒及面骨特徵,但卻被視作一種狒狒。由此可見,這個判據也是模稜兩可的。
2. 寒武紀生物大爆炸(CAMBRIAN EXPLOSION)
寒武紀生物大爆炸大約在五億三千萬年前,在5370萬年左右,幾乎現在多數生物的門同時出現在地球上。從海洋里巨大的管狀蠕蟲,甲殼類到較為高級的脊索動物並存。如果進化確如達爾文所言由點滴的,漸進的方式進行,那么數千萬年的時間無論如何也不足以完成這一歷程。雖然寒武紀之前的多細胞生物化石也有發現,但按進化論的觀點,它們與寒武紀生物並無承傳關係。現代達爾文主義者STEPHEN J GOULD亦無法解釋,稱之為“迷中之迷”(ENIGMA OF ENIGMAS)。
2000年以後的科學界主流意見為:在寒武紀之前的世界其實生命已經很豐富多彩了,多數是類似於今天軟體動物的生命,到了寒武紀早期由於捕食和反捕食的出現,才形成了硬質骨骼、牙齒、甲殼等結構,而這些結構更容易保存,所以看起來像是許多生命突然出現。另外,在對細胞種類的分析方面,結論是雖然寒武紀早期出現了很多種生命,他們的細胞結構、種類複雜程度並沒有大突破,即種類雖多,結構的複雜性和以往及以後的生命相比都是吻合漸變的(《動物的形態複雜性增加》,1994年第2期《古生物學社會》,作者瓦倫蒂諾等。)
3. 人類的起源
的確,從進化論的觀點來看,人類的進化太快了。以人為例,進化論者認為人類誕生的歷程為: 南方古猿(AUSTRALOPITHECUS 200萬年前)---原人(代表為BETICONTUROPS 50萬年前)---- 舊人(代表為NEANDERTHALIAN 10萬年前)---新人(5萬年前)--- 現代人類(5千年前)。在這一過程中,人腦容量以爆炸般的速度增大。而現代人類產生後,進化又仿佛突然消失了,五千年來人的腦容量基本未變,當然這五千年和前面50萬年的變化相比太短了,沒有變化也是正常的。再看腦細胞數量,現代類人猿為10億個,而現代人類約為140億個。單從數字上看是增大了14倍,但智力水平卻發生著跳躍性的變化,主要原因是大腦皮層的進化。
另外,六十年代以來,肯亞OLDUVAI峽谷和RUDOCF湖附近地區的考古發現,也給進化論提出了許多難題。1972年出土了一具類人動物的頭骨化石,編號為KNM-ER-1470,簡稱1470號人。按其分類特徵,它類似於現代人,屬於人屬(HOMO GENUS),比南方古猿和直立猿人(HOMO ERECTUS)要進步得多,但它卻處於290萬年前。1470號人比進化論者所公認的人類始祖南方古猿早九十萬年,比直立猿人早二百萬年。這成為古人類學中的一件懸案,至今沒有能夠歸屬1470號人在進化樹中的位置,很可能是進化的側枝。新研究發現化石的年齡有錯誤,很可能是屬於190萬年前,這樣就能解釋這個難題了。
(三)胚胎學
與比較解剖學類似,胚胎學從生物發育過程中形態的變化,推論出高等動物的胚胎在發育中重現其進化歷程。如人類胎兒在發育之初與豬和鼠的胎兒都具有鰓裂和尾,形態也很相似,其實,這只是表面現象。首先,人類胎兒的“鰓裂”事實上不是鰓裂,而是快速發育的皮膚的褶皺;其次,人類胎兒的尾狀結構也只在初期尾,後期是神經管。在蛙胎髮育過程中的尾狀結構亦是神經管,需要補充的是,最原始的脊索動物如海鞘等,神經管就是在尾部發現的。再次,形態這一表象是由基因決定的。較為普遍接受的HOX基因調控理論 認為,動物形態雖然豐富多彩,但在分子水平上的調控都是復和進化的,因此,動物胚胎髮育之初形態相似,是由於HOX基因調控水平相同而已。這是近年來發育生物學最為重大的發現之一。也就是說,只能通過基因統一性說明進化的同源。
 反進化論
反進化論二. 不可迴避的化學進化
生命產生前在分子水平上的進化(PREBIOLOGICAL EVOLUTION)被稱為化學進化。完整意義上的進化論,不僅要回答生命產生後的生物進化過程,而且還要解決生命物質是如何產生的問題。即怎樣由簡單,無機的小分子進化到複雜,有機的大分子,進而產生生命體。
《DARWIN'S BLACK BOX》一書的作者BEHE教授,對數種有關進化分子生物學的學術期刊(包括JOURNAL OF MOLECULAR EVOLUTION,PROCEEDINGS OF NATIONAL ACADEMY OF SCIENCES等權威雜誌),近十年來所發表的上千篇文章的研究表明,在這方面的進行的工作是零。化學進化被迴避的原因很簡單,這需要上億年的實驗來檢測,目前不可能100%模擬。流行的觀點認為,原始地球擁有還原性大氣,含有氮,氫,硫化氫等成份,不含或含少量的氧。大氣放電經常發生,原始海洋中水溫較高,並有頻繁的地質活動,如地震,火山噴發等。基於這種觀點,1951年STANLEY MILLER進行了著名的“原始湯”(PREBIOTIC SOUP)實驗,他在一個燒瓶中模仿原始地球的環境,用氣體放電模擬雷電。實驗結束後,在產物中分離到了胺基酸,這個結果轟動了科學界。但更多的問題暴露後,樂觀主義漸漸消失了。我們僅舉兩例,說明他們的困難。
(一)化學選擇性
所有生物大分子結構均表現為空間上的有序,蛋白質分子除了特定胺基酸的連線外,還能形成Alpha-螺鏇(HELIX);Beta-摺疊(SHEET)等二級結構;結構域(STRUCTURAL DOMAIN)等三級結構;直至形成四級結構---亞基(SUBUNIT)。雙鏈DNA分子由兩條單鏈組成,除單個核甘酸的連線與氫鍵作用外,還能形成雙螺鏇和超螺鏇(A-DNA;B-DNA;Z-DNA等),所有這些結構都與該大分子的功能緊密相聯。一旦這些結構遭到破壞,該大分子就會失活,所以生物體在合成生物大分子時表現出極高的隨機性。
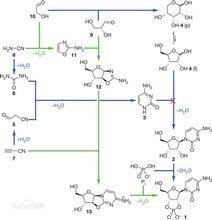
假設第一條具有生命功能的多肽鏈出現在原始海洋中,其序列為A,B,C...(A,B,C...代表天然的人體所需的二十種胺基酸)。其形成的必然條件是:
 化學進化
化學進化(1)A,B,C...等具有足夠的濃度,這樣A,B,C分子才有可能相遇而發生反應。
(2)從胺基酸形成多肽鏈的反應是縮聚反應,每步反應會生成一分子水,從化學平衡的原理看,反應需要脫水劑,否則水將抑制該反應。
(3)假設A與B的結合是隨機的,而結合C時是選擇性的,按熱力學原理,這是一個熵減的反應,必須得有外界能量的輸入。如果以上三個條件全部成立,那么C到底是由什麼因素決定而被選擇的呢?這種問題忽視了熵值定律的前提——封閉系統,而地球本身不是封閉系統,而是有太陽進行能量輸入的。
BEHE教授在談到這個問題時舉了一個例子,他認為,蛋白質絕對隨機,自然地產生,就如同一個人指望把熱水,雞蛋,麵粉,糖和可可粉隨機放在一起加熱一次就能產生一個朱古力蛋糕一樣荒唐。這種觀點早已被科學界拋棄,因為對進化論一知半解的如BEHE等天真地假設進化論是隨機的,如果隨機的話還叫自然選擇嗎?選擇就意味著不是隨機。
關於Behe教授,2007年6月初的《自然》雜誌有詳細的對他的謬論進行駁斥的專題,有興趣的可以去閱讀。
(二)光學選擇性
生物世界是一個不對稱的世界,如果你注意觀察過你的手,你會發現左右手在空間上無法完全重合,但左手和右手的鏡像卻能重合。在化學中這種現象被稱之為“手性”(CHIRALITY),幾乎所有的生物大分子都是手性的。比如,組成人體的糖類一般是右鏇的,而構成蛋白質的胺基酸都是左鏇的。而左鏇的糖和右鏇的胺基酸,幾乎不能被人體利用。一般認為,在非手性的環境中不能產生手性化合物,而只能產生外消鏇體,即等量的右鏇和左鏇體構成的混合物。現代不對稱合成化學通常採用昂貴的手性催化劑,才能使反應按照人所期望的方向進行,得到單一的手性化合物。在MILLER的實驗裡,所有得到的胺基酸都是外消鏇體。假如還用前文所舉的例子A-B-C...序列,又是誰選擇了左鏇的胺基酸呢?眾所周知,最簡單的蛋白質是胰島素--51(肽),那么由自然界隨機地從二十種天然外消鏇體中選擇合成一個51(肽),得到全部由左鏇胺基酸構成,具有生物活性的胰島素分子的幾率有多大呢?簡單的數學計算可以證明是(1/40)的51次方。而在實踐中,其幾率是零,根本就不可能發生。MILLER的實驗帶給進化論者的並不是福音,而是更加劇了其深刻的危機。
