
生物分子的激發態與能量轉移
正文
生物大分子吸收紫外光的能量後,分子內電子躍遷到較高的能級──激發態,激發態電子能量較大、不穩定,容易產生各種變化,其中包括激發能在分子內與分子間的傳遞──能量轉移。葉綠素與視色素等由於受可見光作用,也會產生激發態與能量轉移過程。激發能級的變化 生物大分子吸收紫外光,葉綠素和視色素吸收可見光後,都可出現激發態;只是激發態能級大小有所不同(見圖)。
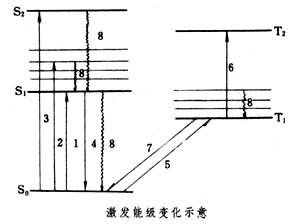 生物分子的激發態與能量轉移
生物分子的激發態與能量轉移蛋白質與核酸的激發態 由於激發態分子可發出螢光或磷光,所以研究激發態往往通過研究發光的特點(如螢光、磷光)以及發光的量子產額、發光的期間(或壽命)、發光的動力學等參量來反映能級的變化和能量關係。蛋白質和多肽的發光主要是由組成蛋白質的胺基酸貢獻的。20種胺基酸中能發光的只有芳香胺基酸,如苯丙氨酸、酪氨酸與色氨酸等。韋伯根據發光特點曾把蛋白質分為A、B兩類。A類蛋白質只含酪氨酸不含色氨酸,它們的發光特點與單純酪氨酸發光相同,說明發光是由蛋白質內的酪氨酸貢獻的。B類蛋白質即含酪氨酸又含色氨酸,它們的發光與單純色氨酸的發光相同,即B類蛋白質發光是由色氨酸貢獻的。近年來又發現了C類蛋白質,這種蛋白質只含苯丙氨酸,發光特點與單純苯丙氨酸相同。B類蛋白質含有酪氨酸和色氨酸兩種芳香胺基酸,而只表現為色氨酸的發光特點,是因為酪氨酸吸收的能量都轉移給色氨酸的緣故。同理A類蛋白質若含有苯丙氨酸,一方面因為苯丙氨酸發光微弱,另一方面它也會轉移能量給酪氨酸,所以A類蛋白質的發光總是體現酪氨酸的特點。各種蛋白質含有這三類蛋質的種類和數量各不相同,但是上述規律卻是相同的。即使象人的血清清蛋白,每一分子含有17個酪氨酸和一個色氨酸,它仍表現為色氨酸螢光的特點。玉米膠原蛋白,因其不含色氨酸,所以它的發光就表現為酪氨酸發光的特點。
蛋白質在室溫下可發螢光,低溫下(液氮)即可發螢光又可發磷光。螢光比磷光的波長短,壽命也短。蛋白質中酪氨酸的量子產額(即發光的量子數與吸收光子的比值)很低,一般少於0.04。比單純酪氨酸的螢光小5~10 倍。這主要是由於蛋白質中的二硫鍵和肽鍵對酪氨酸有猝滅作用。在不同蛋白質中色氨酸的量子產額變動幅度很大 (從0.04~0.6)。對此曾提出過許多不同的機制,但都不能很好地解釋。
蛋白質與胺基酸在極性溶劑中螢光光譜及其峰值波長往往紅移(即向長波方向移動)。紅移是分子激發態的一個普遍現象。如果用光物理的溶劑化模型來解釋,即認為激發分子的電偶極矩往往比基態時大,從而導致激發分子與溶劑殼層中極性分子的強相互作用。若溶劑分子在比螢光壽命還短的期間內能夠重新排列,那末光譜峰值就紅移了。用光化學的激發絡合物模型來解釋,即認為在蛋白質中形成激發絡合物,此時能量降低。而激發絡合物的特徵就是有一個無結構的紅移的螢光光譜。
蛋白質內的色氨酸比單純色氨酸螢光峰值往往藍移,並可由於色氨酸在蛋白質內所處微環境不同而出現發光的異質性。研究蛋白質發光的異質性有助於用發光光譜來探測蛋白質構象的變化及其動力學。
核酸激發態的研究比蛋白質晚10年,其分子較複雜,規律性也較差。核苷酸的最低單線激發態及三重態都是π-π*躍遷。也有人認為在非氫鍵合溶液中單線態是'nπ態,而在氫鍵合溶液中單線是π-π態,三重態在兩種溶液中都是π-π態。核苷酸的螢光量子產額小,系間交換的量子產額也相當小,在室溫下往往猝滅了。幾種核苷酸激發時,它們單線激發態的能級大小的次序是腺嘌呤>尿嘧啶>胸腺嘧啶>鳥嘌呤>胞嘧啶;而三重態的能級大小的次序是胞嘧啶>鳥嘌呤>腺嘌呤>胸腺嘧啶。二核苷酸常具有較寬的無結構的螢光光譜,與單核苷酸相比有較大的紅移。這種譜帶叫做激發二聚體的螢光光譜。多核苷酸在77K下可形成單線態激發二聚體,DNA也如此。DNA 的磷光是來自胸腺嘧啶的三重態。核苷與核苷酸的螢光室溫下與低溫情況相比,光譜加寬、無結構並失去紅移。在室溫下DNA的螢光與鹼基相比光譜加寬有些紅移。在室溫下鹼基的單線態能級次序是腺嘌呤>鳥嘌呤>胞嘧啶>胸腺嘧啶>尿嘧啶。
核苷與酪氨酸的混合溶液的冰凍聚集態,酪氨酸螢光強度下降,核苷則可發螢光,單線態與三重態都如此。但是在少量的色氨酸與腺嘌呤核苷的混合溶液,則後者的螢光猝滅,而色氨酸的螢光則敏化。因為酪氨酸的單線態與三重態能級比所有的核苷都大;色氨酸的三重態能級比所有的核苷能級都小,而它的單線態能級對胸腺嘧啶、胞嘧啶與鳥嘌呤是能量供體,對腺嘌呤和尿嘧啶則是能量受體。
蛋白質與核酸的能量轉移 電子激發能不僅用輻射或內轉換的形式喪失其多餘能量,而且還可用能量轉移方式將能量傳遞給同一分子的其他部分(分子內轉移)或傳遞給另一個相同的或不同的分子(分子間轉移)。因此能量轉移是和電子激發有關的過程。能量轉移有多種形式,最主要的有:共振轉移、電子轉移、絡合物的電荷轉移等。共振轉移指一個激發態分子(供體分子),將激發能以非輻射形式轉移給另一分子(受體分子)的過程。所以共振轉移可以看成是兩個電偶極子之間的電磁偶聯。它們之間的作用,實際是激發的供體對受體部位的電磁擾動的結果。弗爾斯特於1949年提出了著名的共振轉移方程式。即兩個分子間的轉移效率與其距離的六次方成反比。他認為這是發生在單線態與單線態之間的轉移,分子間距離小於100埃。而且能量供體分子的螢光光譜與能量受體的吸收光譜重疊越大,轉移效率也越高。斯特賴斯進一步證明還存在三重態到單線態的能轉移;它本質上是一種延遲螢光,可做為前一種形式的補充。至於三重態-三重態間的能量轉移,則只能發生在距離小於15埃的範圍內。而且它的機制已不是偶極-偶極相互作用,而是一種電子交換作用。斯特賴斯總結了這些結果,提出一個名詞叫“光譜尺”。其意即通過光譜的測定,來測量大分子內生色團間的距離。這是目前能量轉移套用的一個主要方面。另外,利用發螢光物質(螢光探劑)與分子結合,套用螢光方法研究分子構象的變化,其根據之一也是基於能量轉移。
蛋白質中的能量轉移已研究得很多,3種芳香胺基酸的能量轉移次序是苯丙氨酸→酪氨酸→色氨酸。在蛋白質分子內,不同分子(即基團)間的能量轉移已得到公認,但相同分子間的能量轉移,特別是色氨酸→色氨酸之間的能量轉移尚有爭論。
在多核苷酸以及在天然的DNA的同一股上的相鄰鹼基之間也可以發生共振轉移,這是振動弛豫前的單線態能量轉移,轉移率為1012每秒。在DNA中二聚體可直接從激發的單線態形成。在低溫下二核苷酸不論哪一種鹼基吸收光子,都可將其激發成二聚體。相鄰鹼基間可發生單線態的能量轉移,最後轉移到二聚體上。總之,在DNA中可發生如下的一些過程:265納米波長的光可以被A-T或G-C鹼基對同等地吸收,而在振動弛豫尚未發生前,激發能就遷移到相當長的距離(超過4個鹼基對),而優先定位在G-C鹼基對與A-T鹼基對(兩者數量之比為 4:1)。定域在G-C鹼基對的激發,大部分都在單線態能級上猝滅,而定域在A-T鹼基對上的能量則可繼續形成激發絡合物或胸腺嘧啶三重態。所以猝滅與激發絡合物的形成過程都與振動弛豫後轉移強烈地競爭。
生物大分子的能量轉移方式,除共振轉移外,還有激發能借電子和空穴釋放到反應中心的電子轉移、絡合物中單個電子在兩個分子間進行部分轉移的絡合物電荷轉移及溶液中溶質分子吸收能量後將能量轉移到溶劑中的質子轉移等。但通常講的能量轉移,主要是指能量的共振轉移。
